
去甲肾上腺素对腹外侧视前区神经元的作用途径研究
2021-08-04 09:42邴利洁张平平张乐莎王烈成
安徽医科大学学报 2021年7期
邴利洁,张平平,程 娟,张乐莎,王烈成
腹外侧视前区(ventrolateral preoptic nucleus,VLPO)是促进睡眠的中枢,其在睡眠启动中起着重要作用。在中枢神经系统中,去甲肾上腺素(noradrenalin,NA)主要由蓝斑(locus coeruleus,LC)的神经元释放,是觉醒的重要递质。曾有学者提出促睡眠神经元与促觉醒神经元相互抑制的假说,其控制着睡眠觉醒之间的时相转换来维持睡眠或觉醒状态。80% 的 VLPO 神经元为γ-氨基丁酸(GABA)能神经元和甘丙肽能神经元,向脑中与觉醒相关的众多区域发出纤维投射,调节睡眠觉醒时相间的相互转换,是调节睡眠-觉醒节律的关键因素。有研究表明,在VLPO区域中促睡眠神经元约占2/3,中间神经元约占1/3。NA能够兴奋或抑制VLPO中两种类型神经元的兴奋性。有文献表明促睡眠神经元的激活有助于睡眠。然而,目前NA对VLPO区域的这两类神经元的兴奋和抑制作用途径是直接作用途径还是间接作用途径未见报道。该研究利用全细胞脑片膜片钳技术和RT-PCR检测,探究NA对VLPO两种类型神经元的兴奋或抑制作用的途径。
1 材料与方法
1.1 动物与主要试剂
实验动物选用清洁级健康C57BL/6小鼠(15~21 d),购自安徽医科大学实验动物中心,饲养于安徽医科大学基础生理学实验室动物房。饲养环境:室温22~24 ℃,相对湿度40%~60%,人工控制12 h昼夜节律,自由摄食和饮水。主要试剂有:NA购自天津金耀公司,CNQX购自美国abcam公司,AP5和 flumazenil购自美国Sigma公司,逆转录试剂盒购自美国promega公司。1.2 主要仪器
MultiClamp 700B放大器、Axon Digidata 1550A数模转换器(美国Axon公司);振动切片机(德国Leica公司);MODEL P-97电极拉制仪、玻璃电极(德国SUTTER设备公司); 95% O/5% CO的混合气(合肥虹珏公司)。1.3 方法
1
.3
.1
电生理溶液组成 切片液(mmol/L):124 NaCl, 3 KCl, 1.2 NAHPO, 26 NAHCO, 0.5 CaCl, 4 MgSO, 10 dextrose和5 HEPES, 渗透压(798±10)kPa,pH(7.3±0.5), 4 ℃保存。细胞外液(mmol/L):124 NaCl, 3 KCl, 1.2 NAHPO, 26 NAHCO, 2.4 CaCl, 1.3 MgSO, 10 dextrose和5 HEPES,渗透压(798±10)kPa,pH(7.3±0.5),室温使用。电极内液(mmol/L):150 K-gluconate, 5 NaCl, 5 HEPES, 0.4 EGTA, 4 Mg-ATP, 0.5 Tris-GTP和5 phosphocreatine, 渗透压(798±10)kPa,pH(7.2±0.5)。1
.3
.2
玻璃电极拉制 采用电极拉制仪拉制尖端为 1 μm、入液电阻为 3~7 MΩ的玻璃电极。1
.3
.3
VLPO离体脑片制备 从15~21 d的小鼠中制备含有VLPO的切片(厚度300 μm)。小鼠先吸入异氟醚使其麻醉,然后用断头台断头。取脑,放置于含混合气(95% O/5% CO)的4 ℃切片液中,使用振动切片机切割脑片,将切好的脑片移入含混合气的细胞外液中孵育,25 ℃孵育1 h。将切片转移到记录浴槽中,加热细胞外液至30 ℃,进行全细胞膜片钳记录。记录过程中,蠕动泵维持灌流速度为2 ml/min。1
.3
.4
RT-PCR1
.3
.4
.1
VLPO的RNA提取 小鼠吸入异氟醚麻醉后,断头取脑用振动切片机切片,然后对照脑图谱,在VLPO所在的位置,采集含有VLPO的组织。每只小鼠可以取40~50 mg,从6只小鼠中取材200 mg组织,放到1.5 ml EP管中,加入1 ml TRIzol,使用匀浆器吹打研磨。研磨0.5 h后,加入0.2 ml氯仿,振荡30 s,室温放置2 min。12 000 r/min,4 ℃离心15 min。取上清液于新的EP管中,加0.5 ml异丙醇,振荡30 s,室温放置10 min。12 000 r/min,4 ℃离心15 min。弃掉上清液,加入75%乙醇1 ml,振荡30 s,7 500 r/min,4 ℃ 离心5 min。弃上清液,用滤纸小心吸取残留液体,室温干燥30 min,使乙醇充分挥发。沉淀溶于20 μl DEPC水,-20 ℃保存,备用。1
.3
.4
.2
RT-PCR扩增及电泳 Promega GoScript Reverse Transcription System A5000试剂盒说明书对样本RNA进行反转录。PCR扩增程序为:25 ℃退火5 min;42 ℃延伸1 h;4 ℃、5 min;70 ℃延伸15 min;4 ℃保存备用。PCR扩增产物在120 V电压下使用2%琼脂糖凝胶电泳检测。引物由生工生物工程(上海)有限公司设计合成。引物序列如表1所示。
表1 引物序列
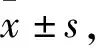
2 结果
2.1 VLPO两种类型神经元的鉴定
高倍显微镜观察下,VLPO中能看到有两种形态的神经元,一种是多极三角形(图1A),这类神经元动作电位的发放频率能被NA抑制(图2A),撤去NA后动作电位发放频率恢复到给NA前水平,是促睡眠神经元;另一种是两极梭形(图1B),这类神经元动作电位的发放频率能被NA兴奋(图2B),撤去NA后动作电位发放频率同样能恢复到给NA前水平,是中间神经元。采用全细胞膜片钳记录VLPO中两种神经元的放电均是自发放电。通过VLPO细胞的不同形态和对NA的不同响应,以此鉴别促睡眠神经元和中间神经元。
图1 小鼠脑片上VLPO中两类不同神经元的形态

图2 VLPO中两种神经元对NA的不同响应
2.2 NA在阻断剂存在下对VLPO促睡眠神经元放电效应的影响
通过显微镜观察,在VLPO中选择钳制形态为多极三角形的细胞进行电生理记录。正常情况下,神经元动作电位的发放是连续的。待神经元的动作电位发放稳定后开始记录,时间记录流程是:稳定记录2 min后,给予NA(100 μmol/L,下同)10 s,观察其抑制效应,然后用细胞外液灌洗2 min,恢复其自身动作电位发放频率,然后给予不同类型阻断剂(antagonist,下同)3 min后,在同时给予(NA+antagonist)10 s,最后用含antagonist的细胞外液灌洗至其动作电位发放频率恢复。该研究把给予NA前20 s作为初始对照(Ctl),给NA时20 s的结果为NA的效应组(NA),细胞外液灌洗1~2 min后的20 s为其恢复组(Wash)。Antagonist灌流3 min左右,即NA+antagonist前20 s为(Ctl+antagonist)组,给予NA+antagonist的20 s为(NA+antagonist)组,1~2 min灌洗后的20 s为(Wash+antagonist)组。Antagonist浓度分别为CNQX(10 μmol/L)、AP5(50 μmol/L)、flumazenil(5 μmol/L),下同。当用NA鉴定细胞为促睡眠神经元时,再进行后续的实验。统计结果如图3所示,在CNQX+AP5存在下,给NA后的动作电位发放频率显著低于Ctl+CNQX+AP5时动作电位发放频率(P
<0.01;n
=8)。在CNQX存在下,给NA后的动作电位发放频率显著低于Ctl+CNQX时动作电位发放频率(P
<0.001;n
=7)。在AP5存在下,给NA后动作电位发放频率显著低于Ctl+AP5时动作电位发放频率(P
<0.001;n
=8)。在flumazenil存在下,给NA后动作电位发放频率显著低于Ctl+ flumazenil时动作电位发放频率(P
<0.05;n
=7)。结果说明,在CNQX、AP5、flumazenil存在的情况下,NA对促睡眠神经元的抑制作用依旧存在。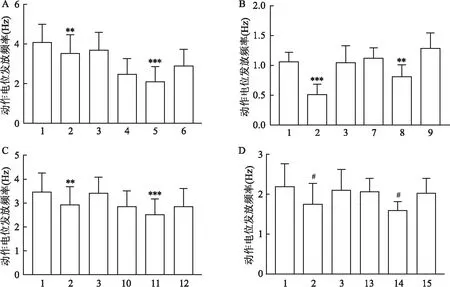
图3 NA在阻断剂存在下对促睡眠神经元动作电位发放频率的效应
2.3 NA在阻断剂存在下对VLPO中间神经元放电效应的影响
通过显微镜观察,在VLPO中选择钳制形态为两极梭形的细胞进行电生理记录。待神经元的动作电位发放稳定后开始记录,时间记录流程和阻断剂使用浓度同2.2。当用NA鉴定细胞为中间神经元时,再进行后续的实验。统计结果如图4所示,在CNQX+AP5存在下,给NA后的动作电位发放频率与Ctl+CNQX+AP5时动作电位发放频率比较,差异无统计学意义(n
=8)。在CNQX存在下,给NA后的动作电位发放频率显著高于Ctl+CNQX时动作电位发放频率(P
<0.01;n
=8)。在AP5存在下,给NA后的动作电位发放频率与Ctl+AP5时动作电位发放频率比较,差异无统计学意义(n
=8)。在flumazenil存在下,给NA后的动作电位发放频率显著高于Ctl+flumazenil时动作电位发放频率(P
<0.05;n
=7)。结果显示,在CNQX、flumazenil存在的情况下,NA对中间神经元的兴奋作用仍旧存在。然而,在AP5存在的情况下,NA对中间神经元的兴奋作用消失。
图4 NA在阻断剂存在下对中间神经元动作电位发放频率的效应
2.4 用不同阻断剂预处理VLPO脑片后分析NA对VLPO神经元两种效应介导的受体途径
通过统计NA对动作电位发放频率抑制率或者兴奋率(%)(NA-Ctl)/Ctl×100%和给予antagonist后,NA对动作电位发放频率抑制率或者兴奋率(%)[(NA+antagonist)-(Ctl+antagonist)]/(Ctl+antagonist)×100%。比较给阻断剂前后NA对VLPO神经元的兴奋或者抑制幅度,结果显示,在CNQX+AP5组中,中间神经元给antagonist后的动作电位发放频率兴奋率(5.223±7.26)%与给antagonist之前的动作电位发放频率兴奋率(40.470±13.59)%差异有统计学意义(P
<0.05,n
=9)。在AP5组,中间神经元给antagonist后的动作电位发放频率兴奋率(3.636±1.71)%与给antagonist之前的动作电位发放频率兴奋率(28.100±6.86)%差异有统计学意义(P
<0.05,n
=8)。而其他组差异无统计学意义。这个结果显示,NA对中间神经元的兴奋作用被AP5阻断。说明NA对VLPO中间神经元的兴奋作用是通过谷氨酸能神经元释放谷氨酸引起的间接效应,该效应由VLPO中间神经元的NMDA受体介导。2.5 VLPO内谷氨酸受体检测
先吸入异氟醚使小鼠麻醉,冰上断头取脑,然后用振动切片机切脑,取出含有VLPO的组织放入1.5 ml的EP管中,经过提取RNA和逆转录的一系列操作,得到的PCR扩增产物在120 V电压下使用2%琼脂糖凝胶电泳检测,结果如图5所示。NMDA受体的3种亚型GluN2A、GluN2B、GluN1在VLPO脑区均有表达。M是DNA marker(DI2000);GAPDH是内参。GluN2A、GluN2B、GluN1是被检测样本。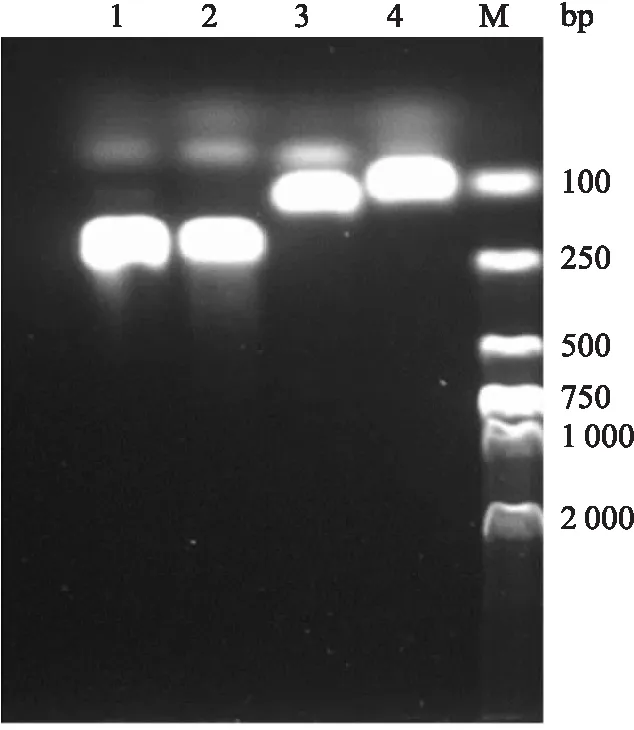
图5 VLPO的RT-PCR检测结果
3 讨论
去甲肾上腺素能神经元聚集的主要核团是LC,研究表明LC在包括注意力、睡眠/觉醒和应激反应在内的许多生理功能中发挥关键作用。实验通过电凝和药理学等方法在LC脑区特异损毁或激活去甲肾上腺素能神经元,提示NA参与了睡眠和清醒行为的调节。Lu et al在他们的研究中表明,在VLPO区域注射顺标示踪物发现,VLPO的轴突对视结节乳头区域神经元的胞体及邻近树突有广泛的投射,对LC、背侧及中缝核、基底前脑有少量的投射。Gallopin et al根据神经元的形态、大小、对药物的响应、递质的组成成功的在VLPO区域鉴定两类神经元,一类是促睡眠神经元,这类神经元的形态呈多级三角形,觉醒一些重要的递质如NA、5-HT对促睡眠神经元的作用是抑制的,同时它们具有低阈值发放特性(low-threshold spike, LTS),并通过单细胞RT-PCR证实了这类神经元是甘丙肽能与 GABA能的。另一类 GABA能神经元,它是两极梭形、NA兴奋性神经元,没有LTS特性。神经生物学研究表明,在中枢神经系统中,GABA在脑中广泛存在,为主要的抑制性神经递质;谷氨酸是中枢神经系统含量最高、分布最广、作用最强的兴奋性神经递质。谷氨酸是种小分子氨基酸,这类神经递质能够结合包括NMDA受体、AMPA受体、红藻氨酸受体的的多个突触后受体。这些受体是阳离子的通道,能使带正电的离子,如NA、K和Ca进入突触后细胞,导致去极化从而激活神经元。因此,在实验设计中,该课题组选取了GABA能受体阻断剂(flumazenil),NMDA受体阻断剂(AP5),AMPA受体阻断剂(CNQX),探究NA对VLPO两类神经元的作用途径。
实验结果显示,NA对VLPO促睡眠神经元的抑制作用均不能被flumazenil、CNQX和AP5阻断。NA对VLPO中间神经元的兴奋作用被AP5阻断,而不能被flumazenil和CNQX阻断。在记录神经元自发放电过程中,任何一种阻断剂还是药物的作用,都会使神经元的动作电位发放频率发生改变,这是因为神经细胞在大脑中接收着各种各样的信号,对神经递质和药物的反应极其敏感。因此,NA对VLPO中间神经元的兴奋作用可能是通过谷氨酸能神经元释放谷氨酸引起的间接效应,该效应由VLPO中间神经元的NMDA受体介导。而谷氨酸神经元的类型和谷氨酸的来源及上游脑区目前还不确定,此类谷氨酸神经元可能来自视网膜的自感光神经节细胞(ipRGCs),因为ipRGCs在脑中有广泛的投射,包括与睡眠相关的VLPO,与非成像视觉密切相关的核团,参与介导生物节律、瞳孔对光反射、睡眠与觉醒周期等生理功能,ipRGCs在大脑中的投射末梢可以释放谷氨酸。
