
度拉糖肽对AD小鼠学习记忆减退和突触退行性变的保护作用*
2021-07-26 06:29姜国晶刘筱涵邓艳秋
医学理论与实践 2021年14期
姜国晶 周 梅 俞 婧 杨 敏 刘筱涵 赵 纲 邓艳秋
1 天津医科大学第二医院老年病科,天津市 300211; 2 天津医科大学基础医学院生理学与病理生理学系;3 天津肿瘤医院病理科
阿尔茨海默病(Alzheimer’s Disease,AD)是以进行性学习记忆力减退为主要临床表现的神经退行性疾病,神经细胞外形成淀粉样斑块(Aβ)、细胞内见神经纤维缠结(NFTs)及神经元、轴突丢失是AD的主要病理表现[1]。神经元突触缺失及其引起的突触可塑性的改变在AD的早期病理变化中占有重要地位,与学习和记忆减退密切相关[2]。葡萄糖代谢异常和胰岛素信号通路受损是2型糖尿病(T2DM)和AD的共同特征,AD也被称为3型糖尿病[3]。胰高血糖素样肽1(GLP-1)作为T2DM药物作用的靶点,受到广泛关注[4]。研究发现GLP-1和GLP-1受体(GLP-1R)广泛表达于中枢神经系统,主要分布在神经元胞体和树突上,GLP-1R基因缺失的小鼠学习记忆能力下降[5]。笔者先前报道长效GLP-1R激动剂度拉糖肽对AD样学习记忆减退的保护作用[6],本实验通过腹腔注射度拉糖肽,联合应用GLP-受体拮抗剂,进一步探究度拉糖肽对散发性AD小鼠的学习记忆、突触蛋白、CREB和Aβ的表达及对神经细胞和突起的影响及其机制。
1 材料与方法
1.1 动物及分组 选取SPF级雄性野生型C57/BL6小鼠32只,9周龄。采用随机数字表法分为对照组(CON组)、模型组(STZ组)、度拉糖肽治疗组(Dul组)和度拉糖肽加抑制剂组(Ex组),每组8只。造模前小鼠常规饲养、适应环境1周。
1.2 实验材料 度拉糖肽(礼来,上海,中国),Ex9-39(中肽,杭州,中国),Streptozotocin (STZ)(Sigma Aldrich,USA),SAP97、SP11抗体(Thermo Fisher Scientific,USA) Aβ1-42、p-CREB抗体、辣根过氧化物酶标记的羊抗小鼠和IgG羊抗兔IgG(Abcam,UK)。无菌度拉糖肽制备:将1.5mg/0.5ml度拉糖肽放于超净台内,取其中0.5ml放入无菌离心管中,4℃无菌条件下保存。腹腔注射时将度拉糖肽与无菌生理盐水混合稀释。
1.3 动物处理和取材 于STZ组、Dul组和Ex组小鼠双侧脑室内注射STZ(3mg/kg体质量),以建立散发性AD模型。注射后21d,Dul组小鼠腹腔注射度拉糖肽,Ex组小鼠注射度拉糖肽和Ex9-39,CON组和STZ组给予等量无菌生理盐水,均持续注射4周。严格遵守《实验动物管理条例》进行实验。水迷宫实验完成后,小鼠断头处死,冰上迅速取脑,取右半脑放入离心管中-80℃冻存用于蛋白免疫印迹实验,左半脑部分固定于4%中性甲醛溶液中用于尼氏染色和高尔基染色。
1.4 学习记忆能力行为测试 自度拉糖肽注射后23d起对小鼠进行学习记忆能力行为学测试,采用Morris水迷宫实验。选取四个象限中点为四个入水点,分别进行以下检测:(1)定位航线实验:将4组小鼠随机从3个不同象限面向池壁放入水中,分别记录60s内小鼠寻找到平台的时间(逃避潜伏期)及其游泳路径。若小鼠于60s内成功找到平台,记录实际逃避潜伏期;若60s内未找到平台,则将逃避潜伏期记为60s,而后研究者手动将小鼠放到平台上并停留30s;每次实验时间为4d,4次/d,路径相同。(2)空间搜索试验:于步骤(1)后次日进行,撤除平台,将各组小鼠分别从第一象限相同入水点面向池壁放入水中,统计小鼠60s内在原平台象限内的停留时间。
1.5 小脑组织SAP97、SP11、CREB和Aβ1-42表达检测 采用Western blot法。取各组小鼠冻存脑组织,冰浴下匀浆,而后4℃、16 000r/min离心10min,取上清液,常规进行SDS-PAGE凝胶电泳,提取蛋白并转移至聚偏二氟乙烯(PVDF)膜,室温下于脱脂牛奶中封闭30min,而后分别加入抗SAP97、SP11、p-CREB、CREB和Aβ一抗,4℃孵育过夜,TBST洗膜,加入辣根过氧化物酶标记的山羊抗小鼠或兔二抗,室温孵育1h,TBST洗膜。将PVDF膜置于扫描仪内,ECL显色。以β-actin为内参。
1.6 尼氏染色 对各组小鼠脑组织切片,脱蜡、复水后,置于尼氏染色液中,于56℃浸染1h。双蒸水清洗,分化液中分化,无水乙醇脱水,二甲苯透明,中性树胶封片,于普通显微镜下观察染色情况并拍照。
1.7 高尔基染色 按试剂盒说明书完成。取2mm厚已固定脑组织,放入3.5%重铬酸钾溶液中1周,每天定时换液。1周后采用双蒸水溶液洗净组织块,放入1.5%AgNO3双蒸水溶液中浸银5d,每天换液1次,常温避光保存。梯度乙醇脱水,二甲苯透明,石蜡包埋,连续切片,每隔5张取1张,厚度为50μm。脱蜡、封片。
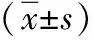
2 结果
2.1 度拉糖肽改善AD小鼠的学习记忆能力 在前4d水迷宫定位航行实验中,与CON组相比,STZ组小鼠的逃避潜伏期增大,Dul组同STZ组小鼠相比,逃避潜伏期缩短,而Ex组小鼠的逃避潜伏期与STZ组的数据接近(图1a)。在空间探索实验,STZ、Ex组小鼠在目标象限的游泳时间比CON组明显减少, Dul组小鼠在目标象限的游泳时间比STZ组小鼠明显增加(图1b)。各组小鼠游泳速度无明显差异,随着训练次数的增加,CON组和Dul组小鼠从盲目的随机式和边缘式寻找平台转变为有目的直线式变化,而STZ和Ex组小鼠轨迹转变不明显(图1c)。这些结果提示度拉糖肽可能通过GLP-1受体改善AD样小鼠的空间学习记忆能力。

图1 小鼠Morris水迷宫实验
2.2 度拉糖肽提高小鼠脑组织神经突触蛋白的表达水平 与CON组相比,STZ、Ex组小鼠脑组织SAP97和SP11表达量下降,与STZ、Ex组相比,Dul组小鼠脑组织两种蛋白的表达升高(图2)。


图2 小鼠脑组织中突触相关蛋白Western Blotting结果
2.3 度拉糖肽促进小鼠转录调节因子CREB的表达,减少Aβ的产生 转录调节因子CREB与调节突触可塑性和学习记忆的形成密切相关,β-actin为内参,同CON组相比, STZ和Ex组小鼠脑组织CREB磷酸化的水平下降,Aβ1-42水平增加。与STZ组相比,Dul组小鼠CREB磷酸化修饰水平上升,Aβ聚集水平下降(P<0.05),提示度拉糖肽可能通过GLP-1受体改善AD样小鼠脑内CREB的活性和减少Aβ聚集(图3)。

图3 小鼠脑组织内CREB蛋白和Aβ的Western Blotting结果
2.4 度拉糖肽改善AD样小鼠脑组织神经细胞的功能活性 CON组小鼠海马CA2区和皮质神经元细胞排列密集、整齐,胞浆中可见丰富尼氏体,呈深蓝色,细胞核蓝染,着色较深;STZ、Ex组小鼠海马CA2区和皮质神经细胞数量减少,细胞浆中尼氏体数量少,呈空泡状,染色较浅;而Dul组小鼠脑组织海马CA2区和皮质神经元细胞尼氏小体比STZ组和Ex组有明显改善(图4)。


图4 小鼠脑组织尼氏染色
2.5 度拉糖肽改善AD样小鼠的神经突起 高尔基染色示,CON组中锥体细胞呈锥体形或三角形,油镜下可见树突和棘突枝干粗大、丰富,二者排列错综复杂;STZ、Ex组树突和棘突断裂,分支减少,枝干纤细;Dul组树突和棘突较STZ组和Ex组明显丰富粗大(图5)。

图5 小鼠大脑皮质神经元的高尔基染色图片
3 讨论
小鼠脑室注射STZ是制备散发性AD动物模型的常用方法。本研究结果显示,STZ处理小鼠的学习和记忆能力较CON组小鼠明显减退,与笔者之前其他研究者的报告一致[7],而度拉糖肽通过GLP-1受体明显改善了模型小鼠的学习记忆能力。脑内GLP-1与GLP-1R结合可激活GLP-1信号,可通过脑—肠—GLP轴抑制食欲,调节神经元轴突生长及海马神经元回路活性、上调酪氨酸羟化酶活性、促进儿茶酚胺生成[8]。外源性给予GLP-1类似物能明显提高脑组织的GLP-1的表达,影响胰岛素信号并激活胰岛素信号分子如cAMP-PKA和PI3K-AKT途径[9],并调节脑内的葡萄糖代谢,AD患者认知能力下降与其脑内胰岛素抵抗、葡萄糖代谢失调等密切相关,GLP-1信号减弱参与认知功能减退和AD神经退行性变,而长效的度拉糖肽可能通过提高胰岛素信号和糖代谢改善模型小鼠的学习记忆能力。
神经元突触缺失和突触可塑性变化是AD早期病理变化,在AD学习和记忆功能障碍发病中起重要作用。突触蛋白是突触可塑性的结构基础,突触相关蛋白97参与控制神经突触受体密度和信号强度,突触小泡蛋白存在于突触前成分的突触囊泡膜上,可间接反映突触的数量、分布和密度,常用作突触密度的标志物[10]。笔者在实验中发现两种蛋白的表达在STZ组明显降低,伴有CREB的活性减低和Aβ1-42的增加,且在尼氏和高尔基染色可以明显观察到尼氏小体的减少和神经突起的减少包括树突缩短和树突棘的明显减少。研究发现:Aβ1-42通过降低树突棘密度,引起神经元损失,进而影响突触可塑性,使与记忆相关的突触功能受损[11]。既往研究[12]发现,早期AD的组织病理学表现为神经元树突棘、突触数量减少,且突触蛋白水平下降早于突触数量减少,进而突触功能受损,降低神经递质传递效率,进而导致Aβ沉积于突触内,产生神经毒性,进一步损伤突触可塑性,最终进展为神经退行性变和认知障碍。动物研究发现,Aβ过度表达可抑制突触蛋白Syn、PSD-95表达。CREB是一种选择性结合CREs 的核蛋白的转录增强因子。主要对PKA等信号发生应答反应。未经磷酸化的CREB主要存在于核内,cAMP激活PKA后,活化PKA进入细胞核,使CREB磷酸化,调节靶基因的转录,这些基因与记忆的形成有关。其中CREB 的133 位丝氨酸残基(Ser133) 对CREB的转录活性起着重要作用[13]。有研究[14]报道,GLP-1类似物利拉鲁肽可促进AKT磷酸化,提高cAMP水平,同时可抑制GSK3β磷酸化及神经细胞凋亡。DPP4抑制剂西格列汀调高GLP-1的水平,提高突触相关蛋白97、突触蛋白-1、突触小泡蛋白和PSD95在3×Tg小鼠脑组织中表达下调,与改善CREB的活性相关[15]。因此,度拉糖肽可能通过提高GLP-1受体调节cAMP、PI3K/AKT等信号分子,影响CREB ser133的磷酸化,活化的CREB进一步改变基因的转录,提高小鼠脑组织突触相关蛋白转录和合成,减少Aβ的沉积,增强突触可塑性,从而达到神经保护的作用。
因此,长效GLP-1R激动剂度拉糖肽可能通过GLP-1受体,调节CREB的活性,减少AD样小鼠脑内Aβ沉积,增加突触蛋白的表达水平和突触的长度,进而改善神经细胞的突触退行性病变,提高小鼠的学习记忆能力,其机制需进一步深入研究。
猜你喜欢
天津大学学报(自然科学与工程技术版)(2022年2期)2022-11-01
西北民族大学学报(自然科学版)(2022年2期)2022-07-06
中国中医药信息杂志(2022年6期)2022-06-27
中国临床解剖学杂志(2022年3期)2022-06-06
河南大学学报(医学版)(2021年1期)2021-11-26
海外星云(2021年6期)2021-10-14
安阳师范学院学报(2021年2期)2021-04-21
商情(2018年9期)2018-03-29
中国医药导报(2017年11期)2017-06-01
中国市场(2017年5期)2017-03-15
