
乙型肝炎病毒S基因不同区域嵌合丙型肝炎病毒中和抗原表位病毒样颗粒实验研究
2021-06-18 09:44王海峰薛飞肖吕海港
陕西医学杂志 2021年6期
舒 放,王海峰,薛飞肖,吕海港
(1.西北大学附属医院 西安市第三医院检验科,陕西 西安 710018;2.西安交通大学附属红会医院麻醉科,陕西 西安 710054)
目前全球丙型肝炎病毒(Hepatitis C virus,HCV)感染者约7100万人,慢性感染导致肝硬化和肝癌,甚至危及生命。近年来因受感染人口老龄化、口服抗病毒疗法广泛应用等原因,全球HCV流行率快速下降[1],但某些国家和地区注射毒品流行的增加在某种程度上部分抵消了这一下降,导致新的HCV感染增加[2-5],仅靠治疗仍不足以实现HCV感染的消除。由于获得治疗的机会有限、费用较高、耐药性、治愈后再感染等情况的存在,迫切需要制定预防HCV感染的有效措施,尤其是研发出广谱有效的疫苗来阻止全球HCV传播[6-7]。现有研究[8-10]表明疫苗在机体细胞免疫及体液免疫反应中能够表现出持久性的抗病毒作用。在控制和预防传染病方面,疫苗仍然是最具成本效益和最成功的干预措施[11]。病毒样颗粒(Virus like particle,VLP)是不含病毒遗传物质颗粒,形态结构与天然病毒相似的能自我装配的病毒空壳,因此不具备感染性,但具有很强的免疫原性和生物学活性,广泛应用于疫苗研究领域[12-13]。本研究利用乙型肝炎病毒(Hepatitis B virus,HBV)S基因具有装配VLP的特性,在其亲水区和氨基端区插入HCV串联中和表位抗原,通过定量测定乙型肝炎表面抗原(HBsAg)比较纯化浓缩后的VLP含量,为后期HCV VLP疫苗研究奠定基础。
1 材料与方法
1.1 材料与试剂 真核载体pCI-neo、重组质粒pCI-HBS、pCI-NEmS、人胚胎肾细胞293T(HEK293T细胞)由唐都医院韦三华教授惠赠;DMEM培养液、胎牛血清(FBS)购于Gibco;Lipofectamine 2000®转染试剂购于Invivogen;重组乙型肝炎疫苗购于GlaxoSmithKline Biologicals;Amicon®Ultracell-15 100K离心过滤器购于Millipore;TaqDNA聚合酶、限制性内切酶AgeⅠ购于Thermo公司。
1.2 串联HCV中和抗原表位合成 根据前期研究选择了一个 HCV E1 基因区保守的中和表位[ITGHRMAWDMMMNWS(氨基酸序列313~327位)]、两个E2区具有广谱交叉活性的中和表位[QLINTNGSWHIN(412~423)和GVPTYSWGENETD(523~535)]以及一个HVR1的模拟表位(ETYVSGGSAARNAYGLTSLFTVGPAQK)。四者之间用 AAY连接,进行PCR扩增构建串联多中和表位序列Em,两端引入AgeⅠ酶切位点ACCGGT。
1.3 串联HCV中和抗原表位嵌合入HBV S基因亲水区 pCI-HBS为亲水区aa127~128位引入AgeⅠ酶切位点的重组质粒,酶切体系中加入pCI-HBS 10 μl,10×缓冲液 0 2 μl,AgeⅠ酶2 μl,ddH2O 6 μl,37 ℃ 4 h进行酶切。电泳后回收胶,纯化至10 μl。将回收纯化的pCI-HBS与Em进行连接,5×缓冲液2 μl,pCI-HBS 2 μl,Em基因5 μl,T4连接酶1 μl,16 ℃过夜。将连接产物转化DH5α感受态,筛选正确克隆,扩增后提取质粒,送测序后保留插入方向正确的质粒,命名为pCI-S1EmS2。
1.4 制备VLP-NEmS和VLP-S1EmS2 HEK293T细胞培养于含10% FBS的DMEM培养液中(双抗)。细胞生长至密度80%时,用脂质体法将pCI-S1EmS2和pCI-NEmS转染至HEK293T细胞,48 h后收集培养上清,2000 r/min离心15 min,除去细胞碎片等杂质,用Amicon®Ultracell-15 100K离心过滤器以4000 g离心15 min,分别命名为VLP-NEmS和VLP-S1EmS2。
1.5 VLP浓缩及纯化 将1.4中VLP-NEmS和VLP-S1EmS2分别铺于9种浓度梯度的蔗糖溶液(20%~60%)上,置于Beckman超速离心机(Rotor SW41)28000 r/min超速离心16 h。从管底依次收集不同浓度层,每个浓度层取10 μl生理盐水稀释至200 μl,在罗氏E601电化学发光分析仪上对每个浓度层HBsAg临界指数(Cut off index,COI)进行测定,收集浓度最高的浓度层。用节流分子量8000~10000的透析袋在20 mmol/L Tris-HCl(8.0)中透析,48 h后将透析好的样品经0.22 μm滤器除菌,用Amicon®Ultracell-15 100K离心过滤器以4000 g离心15 min,得到纯化、浓缩的VLP,-80 ℃冻存。
1.6 VLP定量测定 通过10次独立实验重复1.4、1.5实验步骤,转染HEK293T细胞后收集培养上清,经过浓缩、纯化制备病毒样颗粒VLP-NEmS以及VLP-S1EmS2。电化学发光法对两种VLP进行HBsAg定量检测,VLP-HBS及乙肝疫苗(HBsAg浓度为20×103ng/ml)作为对照。

2 结 果
2.1 HBV S基因及串联中和表位序列Em扩增产物电泳结果 见图1。将HBV S基因及串联多中和表位序列Em扩增产物加样至1%琼脂糖凝胶(0.5 μl/ml溴化乙啶)进行电泳,可见681 bp及255 bp扩增片段,与预期相符。
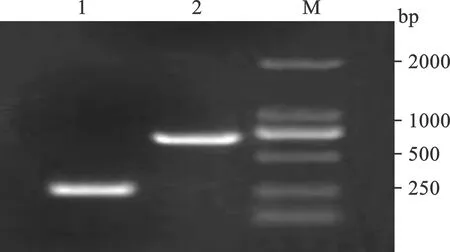
M:DNA Marker 2000;1:HBV S基因扩增产物;2:串联中和表位序列Em扩增产物
2.2 VLP-NEmS和VLP-S1EmS2蔗糖密度梯度离心结果 见图2。分别收集1 ml/管的9层分离片段进行HBsAg定量测定,结果在6号浓度(即45%浓度)蔗糖溶液中收集到的2种VLP浓度最高。
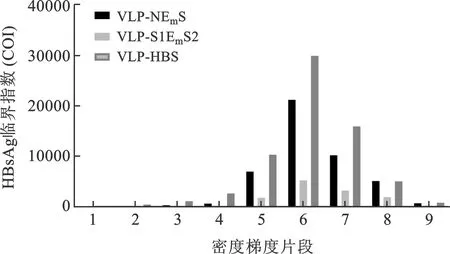
注:1~9号分别代表质量体积百分比浓度为20%、25%、30%、35%、40%、45%、50%、55%、60%蔗糖溶液
2.3 透析、浓缩后VLP浓度比较 见图3。重复转染、纯化、浓缩制备VLP的过程,对10次独立实验的均值结果进行统计学分析。VLP-NEmS、VLP-S1EmS2和非嵌合重组质粒转染的VLP-HBS HBsAg均值分别为(5.83±0.53)×103ng/ml、(1.81±0.76)×103ng/ml和(6.27±0.47)×103ng/ml,其中VLP-S1EmS2低于VLP-HBS(t=16.83,P<0.05),而VLP-NEmS与VLP-HBS均值比较差异无统计学意义(t=1.542,P=0.157)。因此,HCV串联中和抗原表位嵌合于HBV S基因氨基端胞外区形成的病毒样颗粒包装效率高于亲水区。

注:与VLP-HBS比较,*P<0.05
3 讨 论
传统的疫苗研究直到20世纪80年代仍主要基于减毒或灭活病毒,在诱导宿主产生有效T细胞及B细胞免疫应答中显示出良好功效,并能产生持久的免疫力[14]。主要归因于减毒病毒应答能力、表面几何形态重复性、颗粒特性、刺激产生先天性和适应性免疫反应能力等几个关键特性[15],但是传统疫苗在生产效能及对免疫缺陷患者安全性方面有其自身缺陷,因此人们需要寻找新的疫苗研发替代方案。
VLP疫苗具有大多数传统疫苗的特性,由于缺少病毒基因组而无法复制,因此作为疫苗开发的安全性得以保证。大多数VLP外壳由几个相同蛋白质拷贝构成,形成二十面体或螺旋(棒状)结构[16]。绝大多数VLP直径在20~100 nm,这使得其能够自由进入淋巴管到达被膜下淋巴结并选择性地被抗原提呈细胞所摄取[17]。许多病毒结构蛋白都具有自主组装成VLP的能力。VLP可以在170多种不同表达宿主系统中产生,包括细菌、昆虫、酵母或哺乳动物细胞以及植物细胞,某种程度上反映了VLP宿主谱的广泛性。HBV S基因在没有核心蛋白和基因组的参与下能够自我装配,在宿主细胞中产生VLP并分泌至培养上清中,因其可以携带外源性蛋白,易于制备及纯化,能增强外源性蛋白免疫原性,是公认的较好表达载体。使用异源表达系统甚至可以获得三聚体的复杂表位,如人类免疫缺陷病毒(HIV-1型)包膜糖蛋白[18]和流感病毒血凝素[19]的三聚体。
我们根据之前的研究,选取3个HCV抗原表位基因进行串联,分别嵌合于HBV S基因亲水区及氨基端胞外区,构建串联嵌合基因的重组表达载体质粒,转染293T细胞,在培养液上清中分别收集VLP。经过浓缩及纯化,以定量测定HBsAg的方法比较各种VLP浓度,反映HBV S基因不同区域嵌合HCV中和抗原表位表达载体生成VLP的效率。在之前的研究[20]中,我们将不同片段HCV抗原表位分别嵌合在HBV S基因亲水区同一位置,并对其VLP形成情况进行分析,经浓缩、纯化后,几种VLP能够与商品化的乙肝疫苗在HBsAg含量上达到同一数量级,但由于插入片段表位性质及长短不同,收集的几种VLP相比其浓度存在差异,主要表现为HBV S基因亲水区插入的HCV表位序列越长,形成VLP的能力越弱。但理论上嵌合的HCV表位越长,涉及的抗原谱越广,疫苗产生的有效应答概率越大,所以在质和量上产生了矛盾。为了验证插入表位氨基酸数量及位置对包装VLP的影响,我们将3种HCV表位串联形成较长的片段,分别嵌合在HBV S基因亲水区及氨基端胞外区,构建表达载体PCI-NEmS与PCI-S1EmS2,用脂质体法转染293T细胞,在细胞培养上清中收集嵌合病毒样颗粒VLP-NEmS与VLP-S1EmS2,经过多轮重复实验分析嵌合方式对于VLP形成的影响。从HBsAg含量测定结果来看,胞外区嵌合VLP-NEmS与亲水区VLP-S1EmS2浓度结果有统计学差异,胞外区含量较高。
综上所述,HCV中和表位嵌合位置不同造成病毒样颗粒的包装能力差异,嵌合于胞外区氨基端比嵌合于亲水区能够更有效率地包装出病毒样颗粒,成为疫苗研发中可以参考的一种策略。本实验制备的VLP与商品化的乙肝疫苗达到相同数量级,能够满足后续HCV体外培养系统及免疫动物评价中和抗体的实验要求,为诱导广谱交叉保护作用中和抗体的HCV疫苗研究奠定了基础。然而,本研究虽然证明HCV串联中和表位嵌合在HBV S基因氨基端胞外区比亲水区包装出的VLP效率高,但嵌合于HBV S基因不同部位是否能够刺激机体产生相同、有效的中和抗体水平以及交叉保护作用仍需要通过动物模型及中和抗体评价系统做进一步的研究。
猜你喜欢
传染病信息(2022年4期)2022-09-17
中国高原医学与生物学杂志(2022年3期)2022-06-22
橡塑技术与装备(2022年6期)2022-06-02
少男少女·教育管理(2022年3期)2022-05-12
源流(2021年5期)2021-07-30
智慧健康(2021年2期)2021-03-15
文体用品与科技(2019年9期)2019-12-23
温州医科大学学报(2019年4期)2019-04-28
作文周刊·小学一年级版(2016年11期)2016-05-10
汽车电器(2014年5期)2014-02-28
