
Predominant yeasts in Chinese Dong fermented pork (Nanx Wudl) and their aroma-producing properties in fermented sausage condition
2021-06-05 06:29RuifangMiXiChenSuyueXiongBiaoQiJiapengLiXiaolingQiaoWenhuaChenChaoQuShouweiWang
食品科学与人类健康(英文) 2021年2期
Ruifang Mi, Xi Chen,*, Suyue Xiong, Biao Qi, Jiapeng Li, Xiaoling Qiao,Wenhua Chen, Chao Qu, Shouwei Wang,*
a China Meat Research Center, Beijing 100068, China
b Beijing Key Laboratory of Meat Processing Technology, Beijing 100068, China
ABSTRACT
Yeasts are one of the predominant microbial groups in fermented meats. In this study, yeast communities of Chinese Dong fermented pork (Nanx Wudl) were investigated and the technological properties of 73 yeast isolates were evaluated. Through culture-dependent and high-throughput sequencing methods, the main yeast species identified included Pichia membranifaciens, Kazachstania bulderi, Millerozyma farinosa, Candida zeylanoides, Kazachstania exigua, Candida parapsilosis and Saccharomyces cerevisiae. Among these yeasts,P. membranifaciens, M. farinosa, K. exigua and K. bulderi were detected in fermented meats for the first time.A total of 73 yeast isolates was investigated for their lipolytic and proteolytic activities. All yeast species showed lipolytic activity, while proteolytic activity against meat protein was only detected in S. cerevisiae.Assay of aroma-producing potential was performed in a model simulating fermented sausage condition.Inoculation of yeast strains increased volatiles production, especially esters and alcohols. The highest ester production was observed in S. cerevisiae Y70 strain, followed by K. exigua Y12 and K. bulderi Y19.C. zeylanoides Y10 and S. cerevisiae Y70 were the highest producers of benzeneethanol and 3-methyl-1-butanol. S. cerevisiae Y70 with its highest production of branched alcohols and esters could be a promising candidate as aroma enhancer in the manufacture of fermented sausages.
Keywords:
Yeast
Nanx Wudl
Fermented sausage
Aroma
Saccharomyces cerevisiae
1. Introduction
Fermented sausages have been consumed in many countries.Nowadays, the increased awareness of consumers about the health benefits of food has led to the reduction of animal fat and salt in sausages [1]. However, the aroma defect was found in fermented sausages with reduced salt and animal fat [2,3]. In order to enhance the flavor attribute of reformulated fermented sausages, yeast starter cultures with aroma-producing capacity can be used as aroma enhancer [4].
Yeasts are one of the predominant microorganisms during meat fermentation [5]. The processing conditions, ripening time and nature of meat products affect the presence of yeasts [6]. In fermented meats,the yeast species frequently found include Debaryomyces hansenii,Candida zeylanoides, Candida parapsilosis, Yarrowia lipolytica,Rhodotorula mucilaginosa and Saccharomyces cerevisiae [7-9]. The use of yeast could stabilize the typical red color by their oxygenscavenging capability and improve organoleptic characteristics of fermented meats [10]. The enzymatic activities of yeast in fermented meats have been reported by many studies [6,11,12], suggesting that yeasts may contribute to flavor development of fermented meats by their lipolytic and proteolytic activities. Corral et al. [13]reported that lipolysis was accelerated in sodium reduced fermented sausages inoculated with D. hansenii, thus improving the content of free fatty acids, branched aldehydes and esters. S. cerevisiae isolated from salami exhibited proteolytic activity against meat protein and favored the formation of free amino acids which can be transformed into aldehydes, ketones, amines and acids [11]. Yeasts are able to inhibit the lipid oxidation, favoring the formation of esters[14,15]. In addition, yeasts contribute to the inhibition of pathogenic microorganisms, such as ochratoxigenic mould and aflatoxigenic Aspergillus parasiticus in the ecological niche of meats [16,17].
The evaluation of aroma-producing potential of yeasts has been performed previously in different meat models, which are helpful to exclude the influence of other contaminants [8]. The difference in the generation of volatiles by meat-borne yeasts is found not only among species but also within strains from the same species.Andrade et al. [18] found significant disparities of aroma-producing capacity between C. zeylanoides and D. hansenii species. The aromaproducing disparities among different D. hansenii strains were described previously [19,20], suggesting yeasts may enhance and diversify sausage aroma by their specific biochemical activities.
Microorganisms originating from the traditional meat products are promising candidates for meat starter culture because of their good adaptability to the ecological niche of fermented meats [21].Moreover, microorganisms isolated from spontaneous fermentation may offer distinct and attractive aromas based on the specific metabolic activities, improving and diversifying sausage flavor [22].Nanx Wudl, a Chinese fermented meat product, is produced from pork belly without the use of starter cultures. Several studies have revealed the presence of yeast (up to 5.93 lg (CFU/g)) in Nanx Wudl[23]. The yeast genera found in Nanx Wudl were Debaryomyces,Candida, Pichia, Cryptococcus, Torulopsis, Zygosaccharomyces and Trichosporon [24,25]. Therefore, the yeasts isolated from Nanx Wudl could be promising candidates as aroma enhancer for the production of fermented meats. Yet, no survey reported the technological properties of yeasts originating from Nanx Wudl have been published.
In this study, predominant yeasts in Nanx Wudl were identified based on high-throughput sequencing and culture-dependent methods.Based on preliminary screening, 7 strains with strong lipolytic and proteolytic activities were analyzed for their aroma-producing potential in a model resembling fermented sausage condition. The ultimate goal of this study was to obtain new strains with aromaproducing capacity and provide useful information for an appropriate choice of yeast starters.
2. Materials and methods
2.1 Sampling
Twelve Nanx Wudl sampled in this study were collected from three different manufacturers (HF, HJ and HT) in the Dong ethnic minority region of Hunan province, China. These manufacturers are small-scale facilities, producing Nanx Wudl without the use of microbial starters. Main ingredients included pork belly, salt(20-40 g/kg), millet (60-80 g/kg), and additives (sugar and/or Chinese prickly ash).
2.2 High-throughput sequencing and bioinformatics analysis
Sample (25 g) was aseptically taken from each Nanx Wudl sample and homogenized in a sterile bag containing 225 mL physiological saline. The extraction of fungal DNA was conducted using DNeasy Plant Mini Kit (Qiagen, Germany). Amplification was performed using the universal primers ITS1F and ITS1R as described by Bokulich and Mills [26]. Sequencing was conducted using the IonS5TMXL platform (Novogene Bioinformatics Technology, China).The raw reads were merged,filtered and then clustered by UPARSE(v 7.0.1001) software. The OTUs classification was carried out with QIIME software and Unite (v.7.2) reference gene database [27].Chao1 richness and Shannon diversity indices were obtained in R program to evaluate alpha diversity. Principal coordinates analysis(PCoA) was conducted to show the diversity between Nanx Wudl samples based on microbiota. All bioinformatics analyses were performed by Novogene Bioinformatics Technology Company.
2.3 Isolation of yeast strains
2.4 Identification of yeast strains
The purified isolates were subjected to sequencing analysis of ITS and 26S rDNA. Genomic DNA extraction was conducted using DNeasy Plant Mini Kit (Qiagen, Germany). Amplification of ITS region was performed using the universal primer pairs ITS1 and ITS4[28], while 26S rDNA sequence was obtained with the primer pairs NL1 and NL4 [29]. The amplified products were sequenced by Thermo Fisher Scientific Company. By comparing the obtained sequences with type strains in GenBank database, the identification at species level was performed according to a 99%-100% identity criterion.
2.5 Determination of lipolytic and proteolytic activities
Lipolytic activity was determined according to Buzzini and Martini [30] on tributyrin agar. Proteolytic activity was evaluated on PDA agar with skim milk powder (2%) or meat protein (1 mg/mL)extracted from pork as described by Mendoza et al. [6]. After 5 days at 28 °C, the diameter of the clear zone surrounding the colony was measured to indicate proteolytic activity.
2.6 Yeast growth at different temperatures, NaCl concentrations and pHs
Each isolate was propagated in liquid YPD medium for 40 h at 28 °C.The incubated strains were collected by centrifugation at 10 000 g for 15 min using a Sorval LYNX 4000 centrifuge (Thermo Fisher Scientific, Germany), washed twice with sterile physiological saline and re-suspended in the same buffer. Each isolate was inoculated(106CFU/mL) into liquid YPD medium. Effect of different temperatures on yeast growth was observed at 15 and 37 °C. Growth at different NaCl concentrations was detected at 28 °C added with 10% and 15% NaCl. Effect of different pH values on yeast growth was observed at 28 °C in YPD adjusted to pH 3.0 and 5.0. The optical density (OD) of each isolate was determined at 600 nm after 0, 1, 2, 3,4 and 5 days of incubation.
2.7 Yeast growth in fermented sausage model medium
The fermented sausage model-medium was prepared as described by Paik and Lee [31], including 1.2% meat extract(Sigma-Aldrich, USA), 1% glucose, 2% NaCl, 0.2% K2HPO4·3H2O,0.015% MgSO4·7H2O, 0.05% glutamate, and 0.015% NaNO2. After incubation in YPD broth at 28 °C for 40 h, the pellet of each strain was collected by centrifugation at 10 000 g for 15 min using a Sorval LYNX 4000 centrifuge (Thermo Fisher Scientific, Germany). Cells were washed twice with sterile physiological saline and re-suspended in the same buffer. Each strain was inoculated into the model medium(approximately 106CFU/mL) and incubated at 28 °C for 21 days. The model medium without yeast inoculation (non-inoculated control) was incubated under the same conditions.
2.8 E-nose analysis of volatile compounds produced in fermented sausage model medium
The supernatant of incubated culture was separated by centrifugation (5 000 g, 10 min) using a Sorval LYNX 4000 centrifuge(Thermo Fisher Scientific, Germany) and used for electronic nose measurement. The model medium without yeast inoculation was used as control sample. According to Gao et al. [32], a PEN3 electronic nose system (Airsense, Germany) was used to detect different kinds of volatile compounds. The PEN3 electronic nose is equipped with 10 metal oxide sensors. Supernatant sample (1 mL) was added into a 10 mL sample bottle, sealed and heated for 15 min at 45 °C. The measurement time and washing time was 90 and 150 s, respectively.Each sample was measured for 5 times.
2.9 SPME-GC-MS analysis of volatile compounds produced in fermented sausage model medium
The inoculated samples with different aromas compared to non-inoculated control were subjected to SPME-GC-MS analysis.The supernatant from each inoculated sample was separated by centrifugation (5 000 g, 10 min) using a Sorval LYNX 4000 centrifuge(Thermo Fisher Scientific, Germany). Each supernatant sample (3 mL)was placed in 10 mL extraction vial, and 10 μL 48 mg/L 2-octanol(Sigma-Aldrich, USA) was added to each sample as internal standard.The vials were heated at 45 °C for 30 min. Volatiles were collected using a 50/30 μm DVB/CAR/PDMS fibre (Supelco, USA). The fibre was inserted and maintained in headspace of each vial for 30 min at 45 °C. The analysis of volatiles extracted from the supernatants was conducted according to Gutsche et al. [33]. Adsorbed molecules were desorbed in a GC-MS 1310/TSQ8000 system (Thermo Fisher Scientific, USA) at 230 °C for 5 min in splitless mode using helium as carrier gas at a constant velocity of 1.0 mL/min. A DBWAX capillary column (30 m × 0.25 mm × 0.25 μm) was used for separation of the desorbed molecules. The GC oven temperature was held at 40 °C for 3 min, ramped to 50 °C at a rate of 10 °C/min, then heated to 120 °C at 4 °C/min,finally raised to 230 °C at 12 °C/min and kept at 230 °C for 8 min. Mass spectra data were obtained from m/z 35 amu to 400 amu in scan mode with the electron ionization (EI)source at 70 eV. The retention index (RI) was calculated using C5-C20n-alkanes (Sigma-Aldrich, USA) as standards. The identification of volatiles was conducted by matching their RI with those in the mass spectral database (NIST 11). The volatiles were semi quantified using 2-octanol as the internal standard.
2.10 Statistical analysis
Experiments were conducted in triplicate except for electronic nose measurements with 5 replications. The principal component analysis (PCA) was done to analyze the data obtained by electronic nose with WinMuster software. The data obtained by SPME-GC-MS were analyzed using SPSS Statistics 19.0 Core System (International Business Machines, USA). One-way analysis of variance (ANOVA)with the Tukey test was performed to evaluate the effect of yeast inoculation on the production of volatile compounds (P < 0.05).
要尊重学生的主体权利,尽可能多地满足学生的利益诉求,把促进学生的成长和发展作为出发点和目标;要改变教学方式以及师生关系,重新认识学生在教学过程中的地位,要求学生从教学的“边缘”角色转变为教学的参与者,学习变成了教师指导下自主探究的过程。
3. Results and discussion
3.1 Fungal community identified by high-throughput sequencing method
A total of 953 386 raw sequence reads was obtained from 12 Nanx Wudl samples collected, and 934 633 clean reads were collected after quality control process. Good,s coverage had values > 99.90% in all samples, showing that fungal diversity was sufficient with the sampling regime. Fungal richness and diversity were determined by using Chao1 and Shannon indexes as shown in Table 1. The two indexes in samples from manufacturer HT (HT1-HT4) were mostly higher than that in samples from manufacturers HF (HF1-HF4) and HJ (HJ1-HJ4), showing that fungal diversity of HT samples was greater than that of HF and HJ.

Table 1Sequence abundance and fungal diversity in Nanx Wudl.
Fig. 1 depicts the most abundant phyla and genera found in Nanx Wudl samples. Ascomycota, Basidiomycota and Mortierellomycot were the 3 main phyla identified in all samples. The predominant phylum was Ascomycota, which contributed to 70.72%, 98.82% and 73.87% of the fungal sequence reads in HF, HJ and HT samples,respectively. Basidiomycota was the subdominant phylum, which contributed to 3.02% of the total fungal sequences. The dominant fungal genera belonged to the phyla Ascomycota (Pichia, Candida,Kazachstania, Issatchenkia, Millerozyma, Alternaria, Aspergillus,Fusarium of 42.95%, 16.09%, 9.97%, 4.02%, 1.89%, 1.27%, 1.10%,and 0.45%, respectively) and Basidiomycota (Wallemia, Malassezia of 2.23% and 0.54%, respectively). Pichia accounted for more than 35% in most samples, with the exception of HT samples. Candida was dominant in the samples, HT1, HT2, HT3, and HT4, contributing to 29.50%-65.03% of total fungal sequences. The results agreed with other reports that Pichia and Candida are the most common yeast genera in ecological niche of meat [34,35]. However, another frequently encountered yeast genus in meat products, Debaryomyces,was not found in our study. Its absence could be attributed to the specific procedure for production of Nanx Wudl.
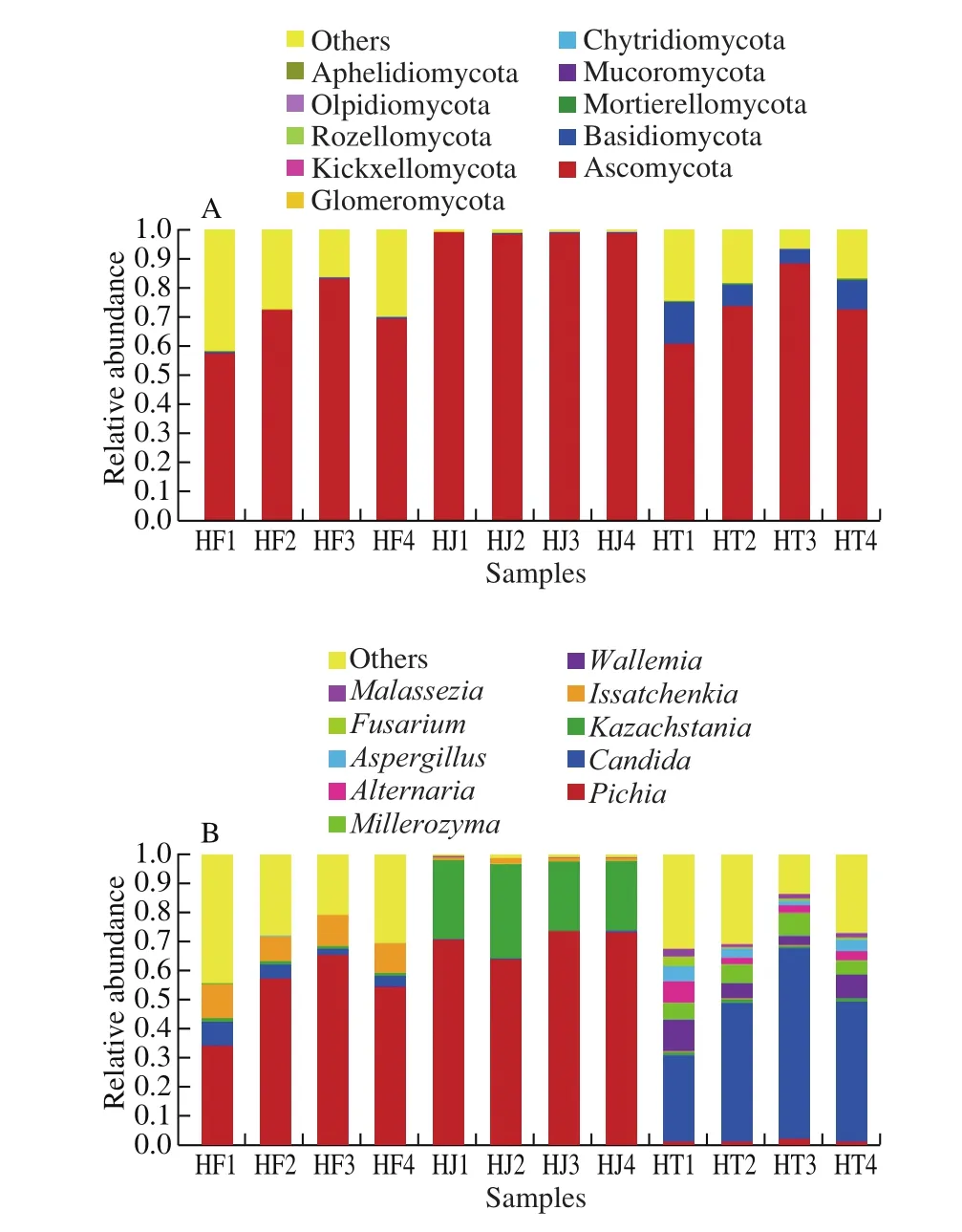
Fig. 1 Relative abundance of fungal community at the phylum level (A) and genus level (B) in Nanx Wudl samples.
The sequences of top 10 yeast species accounted for 36.64%-97.75% of the total sequences. The dominant species includedPichia membranifaciens,Candida metapsilosis,Kazachstania bulderi,Issatchenkia orientalis,Millerozyma farinosa,C. zeylanoides,C. parapsilosis,Kazachstania turicensis,Kazachstania humilis,Candida versatilis.P. membranifaciensaccounted for 33.89%-95.86% in HF and HJ samples.C. metapsilosiswas dominant in the samples, HT1, HT2, HT3, and HT4, contributing to 28.07%-62.46% of total fungal sequences. However, 2.08%-43.70% of the total sequences were not identified at the species level due to the limitation of the reference gene database.
The weighted UniFrac phylogenetic metric was performed to estimate the structural difference of fungal communities among samples (Fig. 2). The principal component 1 (PC1) and PC2 explained 84.24% and 6.56% of the total variance, indicating significant disparities (Pvalues ranged from 0.026 to 0.031) in fungal community between samples from different manufacturers. As expected, samples from the same manufacturer showed similar microflora and clustered closer. The observed disparities in fungal community may be attributed to the initial microbiota composition of the meat and the processing method,which favored the growth of specific microorganism and affected the fungal composition of the final products.

Fig. 2 UniFrac principal coordinate analysis of the fungal community in the Nanx Wudl samples. Red, blue and orange symbols represent Nanx Wudl samples from different manufactures, respectively.
3.2 Predominant yeasts in Nanx Wudl detected by culturedependent approach
The yeast density of each Nanx Wudl sample ranged 4.12-6.30 (lg (CFU/g)). According to colony morphotype, 85 isolates were picked and a total of 73 yeast isolates with representative morphotypes was subjected to genetic analysis based on ITS or 26S rDNA sequence. Table 2 summarized the results of sequencing and NCBI accession.

Table 2Identities (%) of the sequenced genes (ITS or 26S rDNA) of yeast strains and the accession numbers of the entries with the highest identity.

Table 2 (Continued)
The 73 identified yeasts belonged to 7 different species, includingP. membranifaciens,C. zeylanoides,C. Parapsilosis,K. bulderi,K.exigua,M. farinosaandS. cerevisiae. Among them,C. zeylanoidesis one of the predominant yeast species in ecological niche of meat[36]. However, another frequently and abundantly isolated yeast species in fermented meats,D. hansenii, was not found in our study.Other yeast species found in Nanx Wudl includedS. cerevisiaeandC. parapsilosis, which were previously identified from traditional Italian salami [11]. Dorn-In et al. [37] reported the occurrence ofP. membranifaciensin heat-processed meats, but to our knowledge,this yeast species was found in fermented meats for the first time.In addition,M. farinosa,K. exiguaandK. bulderihave not been reported in fermented meat products, although those have been found in other fermented foods, such as sourdoughs and wine [30, 38-40].Therefore,P. membranifaciens,M. farinosa,K. exiguaandK. bulderiwas found in fermented meat products for the first time, suggesting that the specific production conditions may contribute to the selection of specific yeast species during processing.
There were 6 main yeast species identified by both highthroughput sequencing and culture-dependent methods, includingP. membranifaciens,K. bulderi,K.exigua,M. farinosa,C. zeylanoides,andC. parapsilosis. In this study, one predominant yeast species (S. cerevisiae), which was identified by culturedependent approach, was not detected by high-throughput sequencing approach. This could be explained by the preferential detection of yeast species with shorter ITS sequences or more copy numbers of target sequences [41]. In addition, the databases for species annotation should be enriched. Therefore, culture-dependent method contributed to the identification of more yeast species in fermented meats although it was more time-consuming.
3.3 Growth characteristics of yeast isolates
Table 3 depicts the growth ability of 73 isolates at different temperatures (15 and 37 °C), pH values (3.0 and 5.0), and salt concentrations (10% and 15%). The OD600nmvalues were measured to indicate microbial growth. After 3 days of incubation, all the isolates reached maximum OD600nmvalues and then remained more or less constant. The growth disparities of 73 isolates were found in the following 3 conditions: growth at 15 °C, with 10% or 15% NaCl. The numbers of isolates which did not exhibit any detectable growth were as follows: 8 isolates at 15 °C, 31 with 10% NaCl, and 38 with 15% NaCl.

Table 3The technological properties of 73 yeast strains isolated from Nanx Wudl.
As NaCl is one of the essential ingredients during sausage production, the halotolerance of microorganisms should be considered as one selection criteria for starter cultures [42]. In this study,most isolates ofS. cerevisiae,C. parapsilosis,M. farinosaandC. zeylanoidesshowed tolerance to 15% NaCl. Similar results have been reported previously, showing that 85% ofS. cerevisiaestrains originating from salami were able to grow with 15% NaCl [11].In addition, Ozturk [9] reported that 58 strains ofC. zeylanoidesisolated from dry-cured meat product had the capability to tolerate 10% NaCl. In the present study, all isolates obtained could grow at pH 3.0 and 5.0, probably due to the low pHs (< 5.0) of Nanx Wudl samples (data not shown).
3.4 Proteolytic and lipolytic activities of yeast isolates
During meat fermentation, the production of aromas is mainly attributed to proteolysis and lipolysis. Proteolysis influences flavor development through the formation of free amino acids and peptides.Although proteolysis is mainly affected by endogenous enzymes,proteolytic microorganisms also contribute to the hydrolysis of meat proteins during fermentation [43]. In this study, the diameter of the clear zone was measured to indicate the proteolytic activitiy of yeasts.As shown in Table 3, 62 yeast isolates exhibited proteolytic activity agasint milk protein, while only one strain ofS. cerevisiaewas able to hydrolyze meat protein. Similar results have been reported that strains ofC. parapsilosisandC. zeylanoidesoriginating from fermented sausages could not hydrolyze meat protein extracted from beef [6]. However, 15 strains ofS. cerevisiaeoriginating from dry-fermented sausages exhibited the ability to hydrolyze meat proteins by electrophoresis method [11]. These disparities could be explained by the method of evaluation and different sources of yeast. Lipolysis favors the production of free fatty acids which are considered to be important aroma precursors in fermented meats [13]. In this study, most yeast strains (95.89%)exhibited lipolytic activities on tributyrin agar, including 3 strains ofC. zeylanoides(100%). The results were in agreement with Ozturk [9] that mostC. zeylanoidesstrains (91.38%) isolated from dry-cured meat product displayed lipolytic capacity.
The differences in enzymatic activities were discovered not only among species but also within strains from the same species.Some yeast strains showed strong proteolytic and lipolytic activities,indicating that they could probably participate in the production of aromas. It is worth noting thatS. cerevisiaeY70 showed proteolytic activity against meat protein, while high lipolytic activity was also observed in this strain.
3.5 Volatile compounds by different yeasts in fermented sausage model-medium
In fermented sausage model medium, the aromatic capability of the 7 yeast species was evaluated. Based on preliminary screening,7 isolates from different species were selected as they possessed good enzymatic activities and halotolerance, includingC. zeylanoidesY10,P. membranifaciensY11,K. exiguaY12,K. bulderiY19,M. farinosaY39,C. parapsilosisY41 andS. cerevisiaeY70 (Table 4). All yeasts were able to grow in fermented sausage model-medium, and the viable counts of yeasts displayed no significant difference (P> 0.05) between inoculated samples after 21 days of incubation (data not shown).

Table 4The technological properties of selected seven strains for SPME-GC-MS analysis.
The aroma generation potential of different yeast species was initially evaluated by the e-nose method. The principal component 1 (PC1) explained 95.39% of the variance, while PC2 accounted for only 4.26% of the overall variance (Fig. 3). The samples inoculated with different yeast species were divided into 5 clear separated groups based on their aroma profiles. The control sample displayed significant difference in the production of aromas compared to samples inoculated with yeasts, except forM. farinosaY39 andP. membranifaciensY11. The result indicated that the selected yeast strains Y10, Y12, Y19, Y41, and Y70 may participate in the formation of aromas in fermented sausages.

Fig. 3 PCA plot of volatile compounds in fermented sausage model samples inoculated with different yeasts and control batches. Y10, Y11, Y12, Y19,Y39, Y41 and Y70 represented samples incubated with C. zeylanoides,P. membranifaciens, K. exigua, K. bulderi, M. farinosa, C. parapsilosis and S. cerevisiae, respectively.

Fig. 4 Quantification of branched alcohols (A) and ester compounds (B) in fermented sausage model medium inoculated with different yeasts. Control:non-inoculated model-medium; Y10, Y12, Y19, Y41 and Y70: modelmedium inoculated with strains of Y10, Y12, Y19, Y41 and Y70, respectively.Different letters indicate significant differences (P < 0.05) of total branched alcohols or total ester compounds between samples.
The volatile compounds produced by strains Y10, Y12, Y19, Y41,and Y70 were identified by SPME-GC-MS method (Table 5), because these yeasts produced different aromas compared to control. Most of volatiles detected have been previously identified in fermented sausages [44,45]. A total of 43 volatiles were found in control and inoculated samples, including alcohols (10), aldehydes (8), ketones(4), acids (2), esters (9), hydrocarbons (4), sulphur (1) and nitrogen(5) compounds. Table 5 depicts the disparities in the production of volatiles among yeasts and control. Twenty-two volatiles, mostly aldehydes and nitrogen compounds, were found in the non-inoculated control group. These volatile compounds were probably generated through chemical degradation of amino acids and lipids present in the fermented sausage model-medium [8]. As expected, there was no ethanol detected in non-inoculated control, while the production of ethanol was observed in all yeast-inoculated models probably due to microbial metabolism. Other volatiles generated by yeasts in highest abundance were 3-methyl-1-butanol, benzeneethanol and in less abundance esters and 2-methyl-1-propanol. These esters and alcohol compounds were considered as important aroma contributors in fermented sausages. The branched alcohols are responsible for flavors of rose and green grass [3], while ester compounds could afford fruity aromas [46]. It is worth noting that the non-inoculated control had a higher benzaldehyde content than samples inoculated with yeasts.Similar results were reported by Martín [47] and Andrade et al. [18],suggesting that the catalase activity of yeasts may contribute to the degradation of benzaldehyde.

Table 5Volatile compounds detected in fermented sausage model-medium inoculated with different yeasts.

Table 5 (Continued)
Significant disparities (P< 0.05) in the generation of alcohols and esters were observed among samples inoculated with different yeasts(Table 5). As shown in Fig. 4A, all the inoculated samples displayed a significant production of branched alcohols and significant disparities were discovered among yeasts (P< 0.05). Proteolysis favors the production of free amino acids which are considered to be important precursors of 2-methy-1-propanol and 3-methyl-1-butanol [10].The generation of 2-methy-1-propanol and 3-methyl-1-butanol was highest in Y70 strain, probably due to its proteolytic activity against meat protein. Moreover, Y70 was the only strain able to produce 3-methylthio-1-propanol and 1-(2-butoxyethoxy)-ethanol.The production of acid compounds (3-methyl butanoic acid) was only detected in Y70 strain. Regarding aromatic ketones and aldehydes,C. zeylanoidesY10 was the only one that produced 3-hydroxy-2-butanone, while nonanal was only detected inK. exiguaY12.Ester compounds were detected in all inoculated samples exceptC.parapsilosisY41 (Fig. 4B). Lipolytic activity of yeasts could contribute to the production of free fatty acids, thus favoring the generation of ester compounds. During sausage fermentation, coagulase-negative cocci have been associated to the production of ester compounds[48]. However, some studies over the last decade reported the effect of yeasts on the accumulation of free fatty acids through lipolysis,thus improving the content of esters during sausage fermentation[13,15,49]. Nevertheless, the reason for the absence of esters inC. parapsilosisY41 inoculated sample was not clear. Therefore,S. cerevisiaestrains Y70 with its highest production of branched alcohols and esters is a promising candidate as aroma enhancer for fermented sausages. In addition, this is the first time that strains ofK. exiguaandK. bulderihave been reported as ester and branched alcohol aroma contributors in fermented sausage condition.
4. Conclusion
In this study, the technological properties of the yeasts isolated from Nanx Wudl were investigated for the first time.P. membranifaciens,K. bulderi,M. farinosa,C. zeylanoides,K.exigua,S. cerevisiaeandC. parapsilosiswere the 7 main yeast species identified. A total of 73 yeast isolates was investigated for their technological properties. Among the yeast isolates,S. cerevisiaeY70 showed high NaCl tolerance, good lipolytic and proteolytic activities, and the highest production of branched alcohols and esters. Therefore, this strain may offer attractive aroma for fermented sausages. Moreover, the effect of S. cerevisiae Y70 in real dry fermented sausages need to be investigated, as different processing parameters like temperature and formulation composition may affect aroma production of the yeasts.
Conflict of Interest
The authors declare that no conflict of interest exits in the submission of this manuscript.
Acknowledgements
The authors acknowledge the financial support of the National Key R&D Program of China (grant no. 2018YFD0400404).
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
南京理工大学学报(2022年1期)2022-03-17
甘肃教育(2020年18期)2020-10-28
甘肃教育(2020年6期)2020-09-11
甘肃教育(2020年20期)2020-04-13
计算机技术与发展(2017年1期)2017-02-22
电子制作(2016年21期)2016-05-17
华人时刊(2016年13期)2016-04-05
人间(2015年24期)2015-12-26
中国火炬(2009年6期)2009-07-24

- 食品科学与人类健康(英文)的其它文章
- Bioactive compounds and probiotics-a ray of hope in COVID-19 management
- Approaches to evaluate nutrition of minerals in food
- The role of glutamine in supporting gut health and neuropsychiatric factors
- Anti-hyperglycemic effects of dihydromyricetin in streptozotocin-induced diabetic rats
- Aroma profile of two commercial truffle species from Yunnan and Sichuan, China:inter- and intraspecific variability and shared key compounds
- Stability of phenolic compounds and drying characteristics of apple peel as affected by three drying treatments