
园林植物光合节律对夜间光照响应的研究*
2021-04-13 06:59:54刘星滟
重庆工商大学学报(自然科学版) 2021年2期
段 然,刘星滟
(1.重庆工商大学 艺术学院,重庆 40067;2.电子科技大学 光电学院,四川 成都 611731)
0 引 言
园林植物是城市生态系统中的重要组成部分,能够提升城市的艺术品味,也对城市环境的净化等有一系列的促进作用[1]。同时,园林植物也是城市夜景照明中不能移动的有生命载体。统计表明,全球的光污染正以每年 6%的速度递增[1]。城市夜间光环境的肆意塑造扰乱了动植物正常有序的生物节律,甚至影响到城市生态系统的平衡[2]。LED光源以其寿命长、发光效率高、节能环保等一系列优势,逐步替代传统光源,是目前城市夜间光照的主要光源类型。LED光源与天然光光源不同,光源光照强度、光谱能量分布等光学参数均会影响园林植物的光合节律。光照强度过高,园林植物会产生光抑制现象,干扰植物的生长[3];光照强度过低,园林植物为降低自身对光能的需求,呼吸速率和光补偿点也会随之降低[4]。园林植物能够吸收的光源光谱范围在400~700 nm之间[5],光源光谱是促进植物合成有机碳的能源[6],同时也是调节植物生长的信号[7]。植物的光合速率[8]、形态建成等[9]受红光的干扰较大,叶片状态、碳水化合物合成等受蓝光的干扰较大[10-11]。
在植物光合节律的研究中,植物净光合速率是植物光合节律的重要评价指标[12],同时植物的气孔导度[13-14]、蒸腾速率[15]也是植物调节自身与环境平衡的重要生理过程,同样是植物光合节律的重要评价指标。城市夜间光照是基于人眼视看效果的艺术性照明,在夜间利用光照对园林植物进行夜间形象的塑造,未对园林植物的生理生化特征进行考虑,势必对园林植物的光合节律产生影响。在城市夜景照明中,为了营造园林植物夜间艺术效果,多采用白光LED、黄光LED、红光LED、绿光LED及蓝紫光LED对园林植物进行光照[12]。对园林植物的一般性光照强度控制在1 000~4 000 lux之间[12]。目前,针对园林植物光合节律对光照的响应研究内容多是针对天然光的研究,缺乏从园林植物光合节律出发的研究,尤其是关于园林植物光合节律对夜间光照响应的研究。针对夜间光照会对园林植物光合节律造成影响,提出园林植物光合节律对城市夜景照明光照响应的研究,试图得出园林植物光合节律对夜间光照的响应趋势,掌握城市夜间光照对园林环境的影响,为城市夜间光照对城市环境的影响做出预测,具有一定的创新性和实用性。
1 夜间光照与植物光合节律的理论研究
光合作用涉及到植物光能的吸收、能量转换、电子传递、二氧化碳固定等一系列复杂的物理和化学反应过程。当光照强度较低情况下,净光合速率与植物光合有效辐射呈正相关;随着光照强度的增强,净光合速率随植物光合有效辐射的升高而增高,此时净光合速率受温度、二氧化碳浓度、光照强度等因素限制;当光照强度超过一定范围后,净光合速率不再随植物光合有效辐射的升高而增高,净光合速率达到了光合作用的光饱和状态。1935年Baly提出直角双曲线模型公式用来表示植物与光照的关系,表达式为
(1)
其中,Pn为净光合速率;I表示光照强度;α表示植物光响应曲线的初始斜率;Pnmax表示植物最大净光合速率;Rd表示植物暗呼吸速率。对公式(1)进行求导,可得出
(2)
可知,当I=0时,Pn′=α,即当光照强度为0时,得出植物的光响应曲线斜率。由于,Pn′大于0,则式(2)为无极值函数,可见Baly的直角双曲线生物模型为一条没有极值的渐近线,该模型公式不能够准确的表达最大净光合速率Pnmax及饱和光照强度。根据CO2分子在叶片气孔中扩散的物理过程,利用分子扩散及碰撞理论结合流体力学与植物光合生理相关理论,推导得出植物叶片的净光合速率Pn与光照强度的关系为[16]
(3)
其中,Pn为植物叶片的净光合速率;I为光合有效辐射,即光照强度;Rd为暗呼吸效率;α是光响应曲线的初始斜率;β为修正系数;γ为植物叶片光响应曲线的初始斜率与植物最大光合速率之比γ=α/Pmax。该模型能够应用于植物在光饱和点最大光照强度时的光合速率,也可以拟合植物在饱和光强之后光合速率随光强的增加和下降的这一光响应趋势。
通过实验结果拟合,数学模型与Li-6 400光合仪实测光响应拟合曲线相同。故可通过Li-6 400光合仪测试不同光源光谱光照强度下园林植物净光合速率、气孔导度及蒸腾速率,从而掌握园林植物光合节律对城市夜间光照的响应,解决园林植物照明中夜间光照影响园林植物生理的问题,同时有助于掌握夜间光照对园林环境的影响。
2 材料与方法
2.1 实验光源
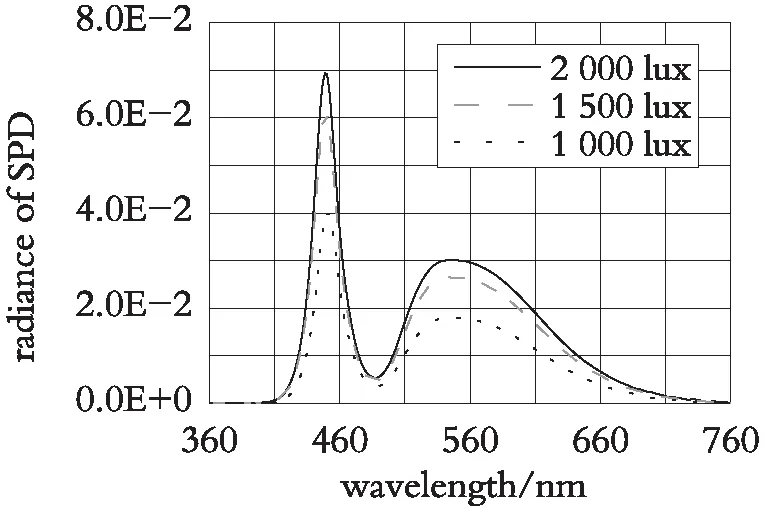
实验选择夜景照明常用的LED光源,白光LED(色温6 500 k)、黄光LED(色温3 000 k)、绿光LED(主波长527.6 nm)、紫光LED(主波长425 nm)、红光LED(主波长618.4 nm),每种光谱光源3盏。所选光源均为某品牌30 W的LED光源,为了降低光源本身产品质量对实验结果的干扰,LED芯片均为某国际品牌。实验前,在光学实验室对光源进行检查并滤光,利用CL-500A分光辐射照度计对光源光谱进行测量。将光源固定通电半小时,稳定后,直接利用CL-500A分光辐射照度计进行光源光谱能量测量,并利用数据管理软件CL-S10W导出光谱能量分布(图1),并得出彩色光源的主波长见表1。
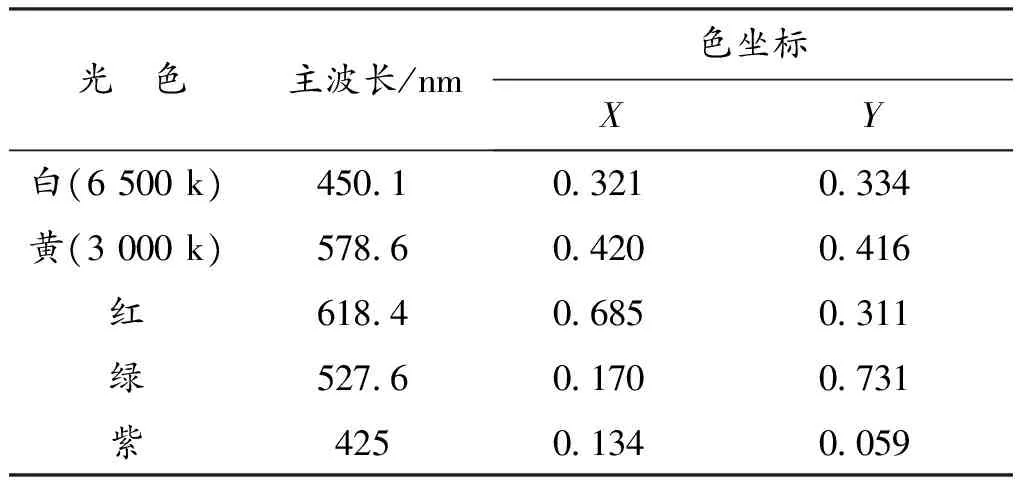
表1 实验光源参数
现实生产的光源产品光谱能量分布与标准光源光谱能量分布有稍许差异,这是产品批量生产不可避免的,所选用的光源灯具误差控制在植物照明可允许范围内,在实验中能够确保光源光谱能量分布与光照强度,满足实验要求(图1(a)—图1(e))。
(a)白光LED
2.1 实验植物
实验地点位于中国重庆(北纬29.35东经106.33)。年平均温度约16~18 ℃,平均日照时数为1 000~14 000 h。实验材料选择窄叶石楠(Photiniaserrulata),蔷薇科、石楠属木本植物,是园林夜景照明中常见的园林植物载体。窄叶石楠叶片色彩不随光照发生变化,可降低对实验的干扰。实验选择5种城市夜间光照光源,每种光谱设置3个光照强度1 000、2 000、3 000 lux,并设1组参照组仅受日光照射不进行夜晚光照,共计16组实验处理,每组处理3株植物,所有植物均生长于标准的花园土壤。按园林照明时间对植物进行夜间光照,照明时间为7∶00 pm~10∶00 pm。照明时利用遮光板遮挡,避免光源相互干扰,日间拆除挡板。
2.3 实验方法
根据植物学实验原理,进行为期 2 a的植物光照实验,选择春季进行园林植物光合节律对夜间光照响应的研究,每 20 d利用利用Li-6 400便携式光合仪(Li-Cor,NE,USA)测量光照实验的园林植物日间和夜间的净光合速率、气孔导度及蒸腾速率数据。日间测量时间为上午9:00,夜间测量时间为下午8:00。日间测量,当显示屏 Ci 为正,且 photo 指标在最大值附近来回跳动时,按下记录键记录一组实验数据。测量选取园林植物顶部新生侧枝的5片叶片,并标记叶片,每片叶重复测量 3 次;夜间测量时,将二氧化碳通气管螺栓调制“scrub”模式,以控制空气湿度,降低夜间由于湿度过高而引起测量数据的不准确。导出光合仪所测得的数据,舍去由于操作错误而造成的错误数据。
3 结果与分析
3.1 园林植物净光合速率对夜间光照的响应
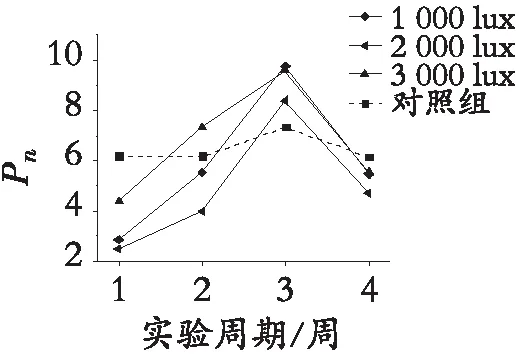
对数据进行整理,并利用origin对数据进行可视化处理,见图2。图中数据为不同光照处理的园林植物日间净光合速率与夜间净光合速率对光照的响应趋势。由于夜间不对参照组植物进行光照,参照组植物夜间无净光合速率数据,故日间测量数据为4组,夜间测量数据为3组。随着夜间光照周期的延长,白光、黄光、红光、绿光、蓝紫光照射的园林植物同时出现日间净光合速率数据逐步提高,夜间净光合速率逐渐下降的趋势。且无论何种光谱的照射,均3 000 lux光照下植物日间、夜间净光合速率数值最高。其中有部分数据出现偏差,但整体趋势一致。园林植物夜间净光合速率对光照的响应强度不断下降,最后趋于平稳;白光、黄光、红光、绿光 2 000 lux 与1 000 lux处理的植物夜间净光合速率值相近,仅蓝紫光2 000 lux下植物净光合速率高于1 000 lux。
白光(日间)
根据图2园林植物净光合速率对夜间光照的响应趋势,可知:只要进行人工光源照射,植物的夜间净光合速率就会大于零;园林植物净光合速率对夜间光照的响应强度低于园林植物对天然光照的响应;参照组园林植物日间净光合速率虽受气候等环境因素干扰,但净光合速率波动变化较小;随实验光照周期的延长,园林植物日间的净光合速率波动非常显著,且变化趋势一致,与所受光照强度、光谱能量分布关联不大;园林植物夜间净光合速率对夜间光照的响应强度随光照周期的延长而降低,最后趋于平稳;第3次数据获取由于天空照度变化,植物日间净光合速率波动大;夜间净光合速率趋于平稳。园林植物夜间净光合速率对光照的响应强度不随日间天空照度变化而发生改变。
3.2 园林植物气孔导度对夜间光照的响应
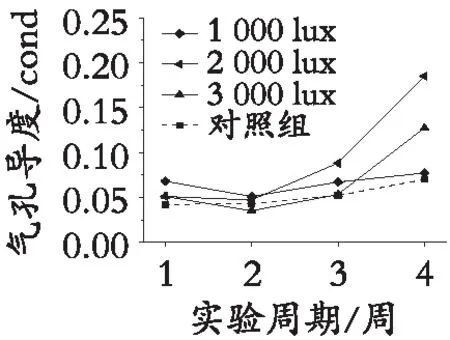
整理测量数据,利用origin对数据进行可视化处理,见图3。图3中数据为不同光照处理的园林植物日间气孔导度与夜间气孔导度对光照的响应趋势。由于夜间不对参照组植物进行光照,参照组植物夜间无气孔导度数据,故日间测量数据为4组,夜间测量数据为3组。随着夜间光照周期的延长,白光、黄光、红光、绿光、蓝紫光照射的园林植物同时出现日间气孔导度数据逐步提高,夜间气孔导度也逐渐上升的趋势。且变化整体趋势大致相同,气孔导度函数斜率变化一致,其中园林植物气孔导度对 2 000 lux白光的响应最强烈;对1 000 lux黄光响应较强。
白光(日间)
根据图3园林植物气孔导度对光照的响应趋势可知:园林植物气孔导度对天然光照的响应强度高于对夜间光照的响应;参照组园林植物仅受日光照射,植物气孔导度曲线较平稳;随实验周期的延长,植物日间的气孔导度波动有明显增强的趋势;植物夜间气孔导度随着光照周期的延长呈现增强趋势。
3.3 园林植物蒸腾速率对城市夜间光照的响应
整理测量数据,利用Origin对数据进行可视化处理,见图4。图4中数据为不同光照处理的园林植物日间蒸腾速率与夜间蒸腾速率对光照的响应趋势。由于夜间不对参照组植物进行光照,参照组植物夜间蒸腾速率数据,故日间测量数据为4组,夜间测量数据为3组。随着夜间光照周期的延长,白光、黄光、红光、绿光、蓝紫光照射的园林植物同时出现日间蒸腾速率先降低后升高最后趋于稳定的趋势,夜间蒸腾速率同时出现先降低后逐渐升高的趋势。且无论何种光谱的照射,趋势相同。
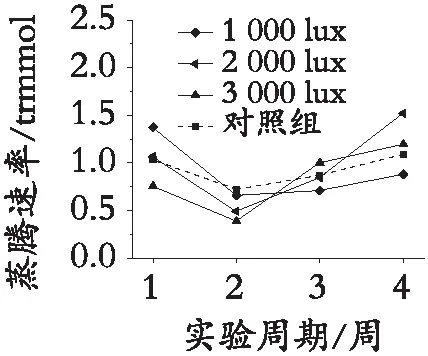
白光(日间)
根据图4园林植物蒸腾速率对光照的响应趋势,可知:园林植物蒸腾速率对天然光的响应整体高于园林植物对夜间光照的响应;参照组园林植物仅受日光照射,园林植物蒸腾速率曲线波动较小;随实验周期的延长,植物日间蒸腾速率波动较大,但最终稳定,且数值趋于不变,变化趋势与植物净光合速率的响应趋势相同;植物夜间蒸腾速率随着光照周期的延长而增强,不同光照强度下蒸腾速率数据的增长速率稍有不同;受光照处理的植物蒸腾速率抗环境干扰能力较差,极易随环境发生变化。
4 结论与讨论
在园林植物光合节律对夜间光照的响应研究中,园林植物的净光合速率、气孔导度及蒸腾速率对夜间光照均有积极的响应,但最终趋于平稳,且最终数值高于未受人工光照前的数值。随光照周期的延长,园林植物不断进行自身的调整,以响应人工光源的光照,最后适应夜晚光照,光合节律数据趋于平稳。在第2次测量时,园林植物光合节律数据发生了波动,所有植物日间所测得的园林植物气孔导度均呈现了低值,分析可知,园林植物在进行夜间光照后,园林植物积极响应夜间的光照,从而不断调节自身光合节律过程,经过较长时间的生理生化调整过程后光合速率趋于平稳,后期继续进行园林植物日间气孔导度测量时,园林植物气孔导度数值逐步上升,园林植物逐渐适应夜间光照情况。第3次测量时,园林植物日间净光合速率数据发生较强烈变化,分析得出,由于天空天然光照度增强,园林植物整体光合指标上升,对当次日间测量的实验数据有所影响,但园林植物夜间光合指标没有发生强烈改变,此时园林植物夜间净光合速率不受日间光合指标变化干扰,也不受日间光照强度的干扰。
园林植物夜间光合节律对光照的响应受光照强度影响极大,光照强度越强园林植物对光照的响应越强。综上可知,夜间只要对园林植物光照,园林植物的光合节律即会发生改变;园林植物光合节律对夜间光照的响应强度随光照周期的延长而降低,最后趋于平稳。紫光、黄光、白光光谱所含的蓝光成分能够促进植物光合效率,此时,二氧化碳经由气孔进入植物叶片细胞,植物的气孔导度被迫提高[17]。气孔导度又直接引发植物蒸腾作用的发生,提高蒸腾速率。虽然植物不能够吸收绿光,但在1 000 lux光照强度绿光LED下,园林植物净光合速率对绿光光谱光照也进行积极响应;2 000 lux及3 000 lux下均是白光、黄光及紫光照射的园林植物光合节律响应最为积极,与先前的研究结果一致[18-19]。得出园林植物光合节律对夜间光照的响应趋势,有助于掌握城市夜间光照对园林环境的影响,同时也补充了绿光、紫光对园林植物影响的规律。
参考文献(References):
[1] GASTON K J,DAVIES T W,BENNIE J,et al.Reducing the Ecological Consequences of Night-time Light-pollution:Options and Developments[J].Journal of Applied Ecology,2012,49(6):1256—1266
[2] 杨春宇,段然,马俊涛.园林照明光源光谱与植物作用关系研究[J].西部人居环境学刊,2015,30(6):24—27
YANG C Y,DUAN R,MA J T.The Study of Garden Lighting Spectrum and Plants[J].Journal of Human Settlements in West China,2015,30(6):24—27(in Chinese)
[3] ZARCO T P J,PUSHNIK J C,DOBROWSKI S.Steady-state Chlorophyll A Fluorescence Detection From Canopy Derivative Reflectance and Double-peak Red-edge Effects[J].Remote Sensing of Environment,2003,84(2):283
[4] REIMUND G,GYOZO G.Non_photochemical Chlorophyll Fluorescence Quenching and Structural Rearrangements Induced by Low pH in Intact Cells of Chlorella Fusca(Chlorophyceae)and Mantoniellasquamata(Prasinophyceae)[J].Photosynthesis Research,2001,67(3):185-190
[5] Plant Factory Laboratory.Light Source for Plant Cultivation[EB/OL].http://www.sasrc.jp/kougen.html.Retrieved 2012-03-20
[6] BROUWER B,ZIOLKOWSKA A,BAGARD M,et al.The Impact of Light Intensity on Shade-induced Leaf Senescence Plant[J].Cell & Environment,2012,35(6):1084—1098
[7] 郝洛西,曹亦潇,汪统岳,等.面向人居健康的城市夜景照明:进展与挑战[J].照明工程学报,2019,30(6):1—6
HAO L X,CAO Y X,WANG T Y,et al.Urban Nightscape Lighting for Human Settlement Health:Progress and Challenges[J].Journal of Illumination Engineering,2019,30(6):1—6(in Chinese)
[8] MATSUDA R,OHASHI-KANEKO K,FUJIWARA K.Photosynthetic Characteristics of Rice Leaves Grown under Red Light with or Without Supplemental Blue Light[J].Plant Cell Physiology,2004,45(12):1870
[9] 邱娟,谭敦炎,樊大勇.准噶尔荒漠早春短命植物的光合特性及生物量分配特点[J].植物生态学报,2007(5):883—891
QIU J,TAN D Y,FAN D Y.Characteristics of Photosynthesis and Biomass Allocation of Spring Ephemerals in the Junggar Desert[J].Plant Evology,2007(5):883—891(in Chinese)
[10] KENDRIK R E,KRONENBORG G H M.Photomorphogenesis in Plants[M].2nd Edition.Dordrecht Netherlands:Kluwer Academic Publishers,1986
[11] MATTSON N S,ERWIN J E.The Impact of Photoperiod and Irradiance on Flowering of Several Herbaceous Ornamentals[J].Scientia Horticulture,2005,104(3):275—292
[12] 段然,杨春宇,苏加福.园林照明对窄叶石楠光合指标的影响[J].同济大学学报(自然科学版),2018,46(7):951—955
DUAN R,YANG C Y,SU J F.Effect of Landscape Lighting on Photosynthetic Indices of Serrulata[J].Journal of Tongji University(Natural Science),2018,46(7):951—955(in Chinese)
[13] NEILK L,LANDRU L,WU J G.Effects of Urbanization on Flowering Phenology in the Metropolitan Phoenix Region of USA Fingdings from Herbarium Records[J].Journal of Arid Enviroments,2010,74(4):440—444
[14] GERRISH G A,MORIN J G,RIVERS T J,et al.Darkness as An Ecological Resource:The Role of Light in Partitioning the Nocturnal Niche[J].Cecologia,2009,160(3):525—536
[15] 王海珍,韩路,徐雅丽,等.不同温度下灰胡杨叶片气孔导度对光强响应的模型分析[J].生态环境学报,2015,24(5):741—748
WANG H Z,HAN L,XU Y L,et al.Model Analysis of the Stomatal Conductance Response to Light in Populus Pruinosa at Different Temperatures in the Taklimakan Desert[J].Ecology and Environmental Sciences,2015,24(5):741—748(in Chinese)
[16] 叶子飘,于强.植物气孔导度的机理模型[J].植物生态学报,2009(4):148—158
YE Z P,YU Q.Mechanism Model of Stomatal Conductance[J].Chinese Journal of Plant Ecology,2009(4):148—158(in Chinese)
[17] 许大全,高伟,阮军.光质对植物生长发育的影响[J].植物生理学报,2015,51(8):1217—1234
XU D Q,GAO W,RUAN J.Effect of Light Quality on Plant Growth and Development[J].Plant physiology,2015,51(8):1217—1234(in Chinese)
[18] 刘晓英.LED光源对樱桃番茄生育和光合作用影响的研究[D].南京:南京农业大学,2010
LIU X Y.Effect of LED Light Source on Growth and Photosynthesis of Cherry Tomato[D].Nanjing:Nanjing Agricultural University,2010(in Chinese)
[19] 周成波,张旭,刘彬彬,等.补光光质对叶用莴苣光合特性的影响[J].植物生理学报,2015,51(12):2255—2262
ZHOU C B,ZHANG X,LIU B B,et al.Effect of Light Quality on Photosynthetic Characteristics of Leaf Lettuce[J].Journal of Plant Physiology,2015,51(12):2255—2262(in Chinese)
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
节水灌溉(2021年5期)2021-05-26 06:49:38
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
中国农业科技导报(2020年12期)2020-03-15 10:50:08
新课程·下旬(2017年7期)2017-08-14 11:12:50
西北林学院学报(2015年5期)2015-01-03 05:12:20
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
植物营养与肥料学报(2011年2期)2011-10-26 03:52:46
