
类胡萝卜素结构及在动植物中的功能与生理活性
2021-04-12 08:41:54任建敏
重庆工商大学学报(自然科学版) 2021年2期
任 建 敏
(重庆工商大学 环境与资源工程学院,重庆 400067)
0 引 言
类胡萝卜素在自然界生物中分布广泛,结构和功能多种多样,是最重要的天然色素之一[1-2],在19世纪初最先被发现存在于红辣椒、藏红花、红木、胡萝卜和秋天落叶。1906年,Zwet等使用柱层析法从绿叶成功分离胡萝卜素、叶黄素和叶绿素。在1930年,Karrer等阐明了β-胡萝卜素和番茄红素的结构,并认知β-胡萝卜素是维生素A的前体。至2018年,约有800多种类胡萝卜素已经从自然界分离鉴定[2]。
类胡萝卜素是迷人的类异戊二烯化合物,可以转化为许多其它化合物,包括类维生素A,在动植物生理过程中起重要作用。
1 类胡萝卜素的结构
类胡萝卜素是四萜烯色素,表现出黄、橙、红和紫色[3],所有植物与一些非光合原核生物、真菌和少数动物,可从头生物合成,但大多数动物与人只能经食物链传递、积累于机体中[4-5]。
类胡萝卜素一般由9个共轭双键多烯链与两端基结合,分为胡萝卜素和叶黄素两大类[4-5]。胡萝卜素是碳氢化合物,如α、β、γ、ε、φ、κ胡萝卜素和番茄红素,自然界约有50多种胡萝卜素。叶黄素是氧化的胡萝卜素,分子中有一个或多个羟基、羰基、甲氧基、环氧和呋喃氧等含氧基团,如β-隐黄质、黄体素、玉米黄质、虾青素、墨角藻黄素和多甲藻素,含氧基团使类胡萝卜素分子结构发生复杂多样的变化,极性的改变易于与机体脂肪酸、糖和蛋白等结合,形成不同功能的活性分子[1]。大多数类胡萝卜素由C40骨架的8个异戊二烯单元构成,有些类胡萝卜素含C45 或C50骨架,称为高类胡萝卜素。碳骨架少于C40的类胡萝卜素,被称为脱辅基类胡萝卜素。脱辅基类胡萝卜素作为C40类胡萝卜素代谢降解产物,约120多种存在于某些植物和动物。类胡萝卜素基本结构与常见胡萝卜素和叶黄素结构如图1所示。
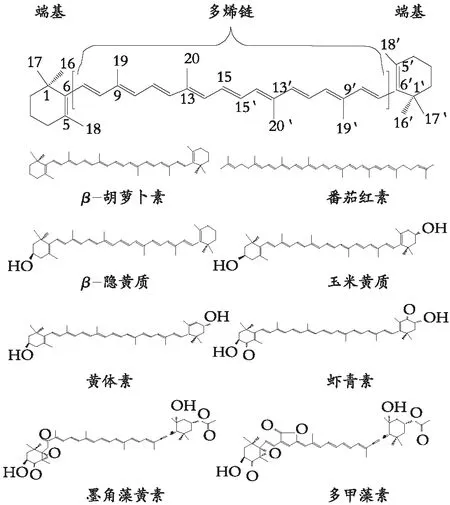
图1 类胡萝卜素的基本结构、末端基与典型的胡萝卜素和叶黄素
2 生物合成
植物与一些非光合原核生物等合成类胡萝卜素,首先由乙酰辅酶A或丙酮酸甲羟戊酸与丙酮酸非甲羟戊酸途径,形成二甲丙烯焦磷酸。后经香叶基焦磷酸与香叶基香叶基焦磷酸,合成类胡萝卜素C40骨架的八氢番茄红素[5]。
八氢番茄红素是含3个共轭双键的无色类胡萝卜素,通过番茄红素脱氢酶逐步脱氢,经六氢番茄红素、ζ-胡萝卜素、链孢红素,形成番茄红素;番茄红素在环化酶作用下,生成如α、β-端基类胡萝卜素,后经胡萝卜素羟化酶、酮醇酶与环氧酶催化,在生物体不同介质与外在条件,生成不同含氧基团的叶黄素[6],如图2所示。
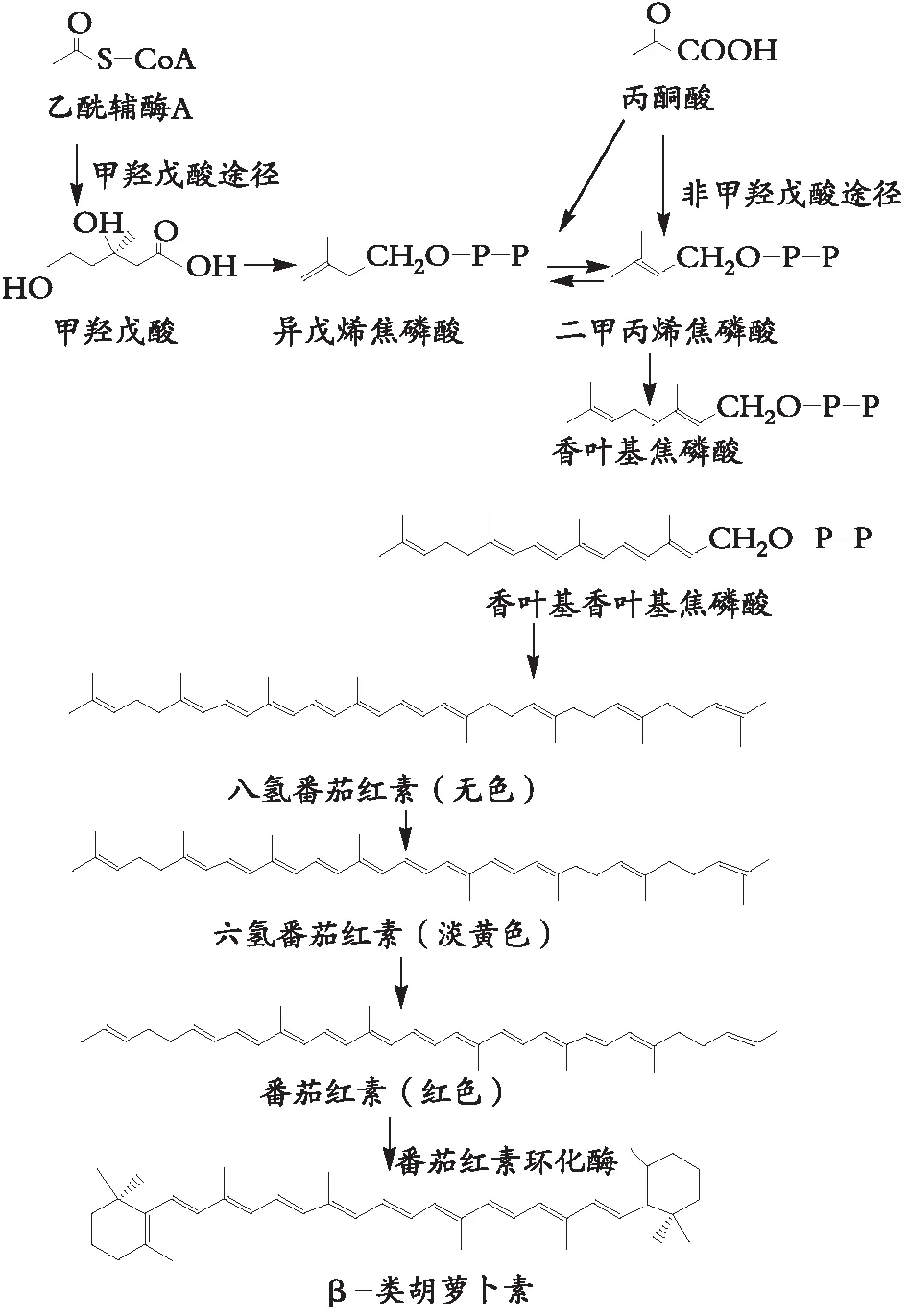
图2 类胡萝卜素的生物合成途径
3 类胡萝卜素的功能与生理活性
3.1 植物中的类胡萝卜素
3.1.1 光合器官
在植物光合器官,类胡萝卜素与叶绿素是参与光合成与光保护,必不可少的色素。光合器官捕光色素天线蛋白吸收光能,传递给叶绿素,形成高能量单线激发态叶绿素,用于反应中心的光化学反应;当光合器官获取光能超过光合反应利用光能时,光能过剩,易引起氧化损伤与光合抑制,甚至机体死亡[6-7]。
3.1.2 非光合器官
类胡萝卜素在非光合器官如植物的果实、果皮、种子、根和花,通过次生代谢反应,如被ROS氧化、多烯链裂解、顺反异构化、与脂肪酸、糖和蛋白键合等,伴随结构与颜色变化[6-11],起光保护剂、抗氧化剂、颜色引诱剂等作用[2]。
类胡萝卜素也参与细胞信号传导,产生脱落酸、独脚金内酯和胡萝卜内脂等植物激素,调节植物生长、种子休眠、胚胎成熟和萌发、细胞分裂和伸长等[12]。植物在温度、光颜色和强度、土壤成分、氧化压力等应激响应下,类胡萝卜素也是茶叶等合成重要致香物质的前体[13-14]。一些果实、果皮、种子、树叶在不同季节呈现不同的颜色,引诱鸟类捕食并被传播的红色种子[2,15],都与类胡萝卜素化学结构随介质与外在条件,发生复杂变化紧密相关。
3.2 动物中的类胡萝卜素
与光合生物光合作用产生ROS一样,动物与人呼吸代谢也会持续产生ROS,低浓度是细胞参与生命活动重要的信号分子,但高浓度对细胞脂质、蛋白质和DNA等生物大分子产生氧化损伤,导致慢性炎症。慢性炎症被证实是动物与人类疾病包括癌症、肺部疾病和自身免疫疾病等引起的主要原因[16]。
具有至少一个未被取代的β-环类胡萝卜素,如β-胡萝卜素、β-隐黄质与玉米黄质,被称为维生素A原[17],动物与人只能通过食物摄入或类胡萝卜素氧化裂解生成维生素A,它是动物与人不可或缺的营养素[18]。
大多数动物与人从食物链传递或代谢转化获得的类胡萝卜素,在维生素A生成、基因表达、细胞增殖和分化调控、细胞信号传导、上调抗氧化酶和免疫等方面有重要作用[19-20]。
3.2.1 水生动物
水生动物从食物如藻类和其他动物获取类胡萝卜素,主要有β-胡萝卜素、墨角藻黄素、多甲藻素、硅藻黄素、异黄素和虾青素及代谢物(2,21)。
双壳类(牡蛎、蛤、扇贝、贻贝和柜壳)和被囊动物(海鞘)是滤食动物。它们以微藻类如硅藻、甲藻、蓝藻、绿藻为食,并从中获取类胡萝卜素。硅藻中类胡萝卜素主要是墨角藻黄素,其多个官能团如累积二烯烃、环氧、羰基、乙酰基,经代谢转化呈现多样性结构(21-22)。
多甲藻素含C37碳骨架(图1),是鞭毛藻类重要的红色类胡萝卜素。多甲藻素的累积二烯烃、环氧和内酯环,也易代谢转化(21-22)。
虾青素是甲壳类动物(虾、蟹)的一种特征性海洋类胡萝卜素。许多甲壳类动物摄取膳食藻类的β-胡萝卜素,通过海胆烯酮、3-羟基海胆烯酮、斑蝥黄质、金盏花红素,合成虾青素[2,22]。鲤科鲤鱼、鲫鱼和金鱼,能将玉米黄质转换成虾青素。一些不能从类胡萝卜素如β-胡萝卜素和玉米黄质合成虾青素的海洋鱼类如红鲷鱼、鳕鱼、金枪鱼、黄尾鱼和鲑科鱼类(鲑鱼和鳟鱼),只能从食物链中获取虾青素。几种海洋鱼类的鳍和皮肤呈亮黄颜色,是由于虾青素经玉米黄质代谢,生成的胡萝卜二醇所致(21-22)。
许多海洋无脊椎动物如甲壳类动物,将摄入体内的β-胡萝卜素转化为虾青素,积累在表皮、甲壳、卵和卵巢,颜色由β-胡萝卜素黄色变成虾青素红色(11)。海洋无脊椎动物的虾青素,与蛋白结合呈现红色、蓝色或紫色。在海底光条件下,海洋动物用这些颜色伪装,或作为光感受器,或对光可能的伤害提供防范。
海洋动物对食物类胡萝卜素转化并积累在不同器官,提高了其抗氧化和光保护活性。如虾青素猝灭1O2、清除·OH、抑制脂质过氧化作用,强于β-胡萝卜素[23],其颜色更适应环境变化。
3.2.2 鸟 类
鸟类从食物链获取类胡萝卜素,其羽毛大部分呈亮红色、橙色和黄色。红色羽毛,至少含10种类胡萝卜素,羽毛鲜艳的颜色是营养良好与健康状况的重要信号,有利于吸引异性促进交配。操控雄性斑马雀食物类胡萝卜素,可唤起对应的细胞介导免疫功能变化和性吸引力[11,24]。
3.2.3 陆地动物
陆地动物机体内类胡萝卜素主要有β-胡萝卜素、β-隐黄质、玉米黄质和黄体素及代谢物[6,11],大多来源于植物性食物。
昆虫是陆地上最多样的动物。蚜虫、粉虱通过从真菌或共生细菌基因转移,获取生物合成基因,可从头合成类胡萝卜素[25]。蓝绿色蚜虫使用共生细菌基因,合成类胡萝卜素和奎宁[26],用于环境伪装与保护着色。甲虫和蜻蜓通过蚜虫类食物链,在体内积累与代谢类胡萝卜素。
棒虫不同季节呈现不同颜色,是体内积累的类胡萝卜素因环境等因素引起不同代谢,产生的结构变化[27]。双斑叶螨为适应过冬漫长的夜晚和更低温度,雌性螨虫进入兼性滞育,体色从微弱的黄色到明亮的橙色,源于体内代谢积累的酮基类胡萝卜素,如3-羟基海胆烯酮、金盏花红素与虾青素等。
哺乳动物从食物链吸收类胡萝卜素能力差异大,人与猴子为更好地适应生存,可同时吸收并积累胡萝卜素和叶黄素[11],在血液和大脑优先积累β-隐黄质[28]。
人体类胡萝卜素主要来自水果和蔬菜,也通过食物链包括家畜、家禽与海洋动物(如牡蛎、蛤、扇贝、贻贝、海胆性腺、鲑鱼和虹鳟鱼)等获取[29]。人类食物链大约50种多类胡萝卜素,在血液中主要有β-胡萝卜素、α-胡萝卜素、番茄红素、β-隐黄质、黄体素和玉米黄质,占类胡萝卜素90%以上[28-29]。在人血浆中也发现番茄红素、黄体素和玉米黄质的代谢物[30-31]。
一些动物类胡萝卜素不同颜色,还用于种内信号(性信号、社会地位与关系信号)和种间(物种、模仿和保护色)识别[11,29]。
4 结 论
类胡萝卜素是自然界所有光合生物与动物包括人所需色素与营养素,具有强大的抗氧化、光保护、着色等功能与生理活性,在维持各生物体持续健康的生命活动中,扮演非常重要的角色。随着科学技术的进步与人们对类胡萝卜素认识的不断提高,资源丰富的类胡萝卜素如番茄红素、β-隐黄质、黄体素、虾青素和玉米黄质等,必将更加广泛应用于食品、保健、护肤与化妆、医药、饲养等行业,以提高人类生活质量与促进人体健康,造福于人类。
参考文献(References):
[1] 姜建国,王飞,陈倩,等.类胡萝卜素功效与生物技术[M].北京:化学工业出版社,2008
JIANG J G,WANG F,CHEN Q,et al.Carotenoid Efficacy and Biotechnology[M].Beijing:Chemical Industry Press,2008(in Chinese)
[2] MAOKA T.Recent Progress in Structural Studies of Carotenoids in Animals and Plants[J].Arch Biochem Biophys,2009,483(10):191—195
[3] KAULMANN A,BOHN T.Carotenoids,Inflammation,and Oxidative Stress-implications of Cellular Signaling Pathways and Relation to Chronic Disease Prevention[J].Nutr Res,2014,34(2):907—929
[4] MELO V L D,LEERMAKERS E,DARWEESH S,et al.The Effects of Lutein on Respiratory Health Across the Life Course:A Systematic Review[J].Clin Nutr ESPEN,2016(13):1—7
[5] ZIELINSKA MA,WESOLOWSKA A,PAWLUS B,et al.Health Effects of Carotenoids during Pregnancy and Lactation[J].Nutrients,2017,9(8):838—862
[6] BRITTON G,LIAAEN-JENSEN S,PFANDER H.Carotenoids Volume 3:Biosynthesis and Metabolism[M].Birkhäser:Basel,Switzerland,1998
[7] 程明,李志强,姜闯道,等.青稞的光合特性及光破坏防御机制[J].作物学报,2008,34(10):1805—1811
CHENG M,LI Z Q,JIANG C D,et al.Photosynthetic Characteristics and Light Damage Defense Mechanism of Highland Barley[J].Acta Agronomy Sinica,2008,34(10):1805 —1811(in Chinese)
[8] PINNOLA A,DALL O L,GEROTTO C.Zeaxanthin Binds to Light-Harvesting Complex Stress-Related Protein to Enhance Nonphotochemical Quenching in Physcomitrella Patens[J].The Plant Cell Online,2013,25(9):3519—3534
[9] DALL O L,CAFFARRI S,BASSI R.A Mechanism of Nonphotochemical Energy Dissipation Independent from Psbs Revealed by A Conformational Change in the Antenna Protein CP26[J].The Plant Cell Online,2005,17(4):1217—1232
[10] CALIANDRO R,NAGEL K A,KASTENHOLZ B.Effects of Altered α-and β-branch Carotenoid Biosynthesis on Photoprotection and Whole-plant Acclimation of Arabidopsis to Photo-oxidative Stress[J].Plant Cell & Environment,2013,36(2):438—453
[11] BRITTON G,LIAAEN-JENSEN S,PFANDER H.Carotenoids Volume 4:Natural Functions[M].Birkhser:Basel Switzerland,2008
[12] TSUCHIYA Y,MCCOURT P,LOHMANN J U,et al.Strigolactones:A New Hormone with a Past[J].Curr Opin Plant Biol,2009,12(5):556—561
[13] 曾晓雄.茶叶中类胡萝卜素的氧化降解及其与茶叶品质的关系[J].茶叶通讯,1992(1):31—33
ZENG X X.Oxidative Degradation of Carotenoids in Tea and Its Relationship with Tea Quality[J].Tea Newsletter,1992(1):31—33(in Chinese)
[14] 宛晓春,夏涛.茶树次生代谢[M].北京:科学出版社,2015
YUAN X C,XIA T.Secondary Metabolism of Tea[M].Beijing:Science Press,2015(in Chinese)
[15] FUJIWARA Y,HASHIMOTO K,MANABE K,et al.Structures of Tobiraxanthins A1,A2,A3,B,C and D,New Carotenoids from Seeds of Pittosporum Tobira[J].Tetrahedron Lett,2002,43(11):4385—4388
[16] HUSSAIN S P,HARRIS C C.Inflammation and Cancer:An Ancient Link with Novel Potentials[J].Int J Cancer,2007,121(11):2373—2380
[17] WEBER D,GRUNE T.The Contribution ofβ-carotene to Vitamin A Supply of Humans[J].Molecular Nutrition&Food Research,2012,56(2):251—258
[18] 江载芳,申昆玲,沈颖,等.诸福棠实用儿科学(上册)[M].第8版.北京:人民卫生出版社,2013
JIANG Z F,SHEN K L,SHEN Y,et al.Zhufutang Practical Pediatrics[M].8th Edition,Beijing:People’s Medical Publishing House,2013(in Chinese)
[19] KARADAS F,PAPPAS A C,SURAI P F,et al.Embryonic Development within Carotenoid-enriched Eggs Influences the Post-hatch Carotenoid Status of the Chicken[J].Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2005,141(2):244—251
[20] 吴灿杰.β-隐黄质的抑癌与免疫增强作用分子机制研究[D].武汉:华中农业大学,2013
WU S J.Molecular Mechanism of Anti-tumor and Immune-enhancing Effects of β-cryptoxanthin[D].Wuhan:Huazhong Agricultural University,2013(in Chinese)
[21] MAOKA T.Carotenoids in Marine Animals[J].Mar Drugs,2011(9):278—293
[22] MATSUNO T.Aquatic Animal Carotenoids[J].Fish Sci,2001,67(5):771—789
[23] MAOKA T,NISHINO A,YASUI H,et al.Antioxidative Activity of Mytiloxanthin:A Metabolite of Fucoxanthin in Shellfish and Tunicates[J].Marine Drugs,2016,14(5):93—1470
[24] BLOUNT J D,Metcalfe N B,Brikhead T R,et al.Carotenoid Modulation of Immune Function and Sexual Attractiveness in Zebra Finches[J].Science,2003,300(5616):125—127
[25] MORAN N A,JARVIK T.Lateral Transfer of Genes from Fungi Underlies Carotenoid Production in Aphids[J].Science,2010,328(4):624—627
[26] TSUCHIDA T,KOGA R,HORIKAWA M,et al.Symbioticbacterium Modifies Aphid body Color[J].Science,2010,330(11):1102—1104
[27] MATSUNO T,MAOKA T,TORIIMINAMI Y.Carotenoids in the Japanese Stick Insect Neophirosea Japonica[J].Comp Biochem Physiol 1990,95(2):583—587
[28] NISHINO A,ICHIHARA T,TAKAHA T,et al.Accumulation of Paptika Carotenoid in Human Plasma and Erythrocytes[J].J Oleo Sci,2015,64(12):1135—1142
[29] BRITTON G,LIAAEN-JENSEN S,PFANDER H.Carotenoids Volume 5:Nutrition and Health[M].Birkhser:Basel,Switzerland,2009
[30] KHACHIK F,BEECHER G R,GOLI M B,et al.Separation and Identification of Carotenoids and Their Oxidation Products in the Extracts of Human Plasma[J].Anal Chem,1992,64(7):2111—2122
[31] KHACHIK F,PFANDER H,TRABER B.Proposed Mechanism for the Formation of Synthetic and Naturally Occurring Metabolites of Lycopene in Tomato Products and Human Serum[J].J Agric Food Chem,1998,46(10):4885—4890
[32] WINGERATH T,SIES H,STAHL W.Xanthophyll Esters in Human Skin[J].Arch Biochem Biophys,1998,355(1):271—274
[33] HEINEN M M,HUGHES M C,IBIEBELE T I,et al.Intake of Antioxidant Nutrients and the Risk of Skin Cancer[J].Eur J Cancer,2007,43(10):2707—2716
[34] ALVES R A,SHAO A.The Science Behind Lutein[J].Toxicology Letters,2004,150(11):57—83
[35] NWACHUKWU I D,UDENIGWE C C,ALUKO R E.Lutein and Zeaxanthin:Production Technology Bioavailability Mechanisms of Action Visual Function and Health Claim Status[J].Trends in Food Science & Technology,2016,49(2):74—84
猜你喜欢
儿童故事画报·自然探秘(2022年3期)2022-04-27 00:17:10
南方水产科学(2022年2期)2022-04-22 02:10:32
食品工业(2021年11期)2021-11-25 09:33:34
当代水产(2021年6期)2021-08-13 08:17:10
落叶果树(2021年6期)2021-02-12 01:29:04
小学生导刊(2017年22期)2017-07-19 13:41:46
小布老虎(2016年18期)2016-12-01 05:47:29
食品研究与开发(2016年16期)2016-04-10 14:35:21
食品科学(2013年17期)2013-03-11 18:27:10
食品科学(2013年8期)2013-03-11 18:21:23
