
胍基乙酸的代谢途径及其在动物生产中的应用
2021-03-31 00:18王子苑舒健虹袁暘暘王小利
贵州农业科学 2021年1期
王子苑,舒健虹,袁暘暘,王小利
(贵州省农业科学院 草业研究所,贵州 贵阳 550006)
0 引言
胍基乙酸(Guanidine acetic acid, GAA)又称胍乙酸、N-咪基甘氨酸(N-Amidinoglycine),是一种氨基酸类似物,在脊椎动物体内经甲基化形成肌酸(Creatine, Cr),并且是合成肌酸的唯一前体物[1]。1835年CHEVREUL首次在骨骼肌中发现肌酸,之后的研究发现动物本身可内源性合成机体所需肌酸的60%,但每天会有1.7%左右的肌酸-磷酸肌酸体系转化成肌酐随尿液排出体外[2],因此机体必须持续地补充肌酸以满足生长需求。营养学家发现,在畜禽饲料中外源补充肌酸能提高动物生长性能,促进肌肉中相关基因的表达以及蛋白质的合成[3-4],然而补充的肌酸稳定性较差、利用率低,因此现在动物生产中多采用胍基乙酸替代肌酸,其已经被证实在提高组织肌酸负荷方面比肌酸更有效[5]。自2009年欧盟正式批准胍基乙酸作为饲料添加剂应用于肉仔鸡以来,胍基乙酸作为一种新型绿色的饲料添加剂受到越来越多动物营养学家的关注。综述了胍基乙酸的代谢途径及其在动物生产中的作用效果,以期为胍基乙酸的进一步研究提供参考。
1 胍基乙酸的来源及代谢途径
WEBER最早从人和狗的尿液中分离出胍基乙酸,并提出胍基乙酸是合成肌酸的前体物质。肌酸磷酸化后生成磷酸肌酸,二者共同组成磷酸原系统,当机体ATP供应不足时,磷酸肌酸在肌酸激酶(Creatine kinase, CK)的作用下将其磷酸基转移到ADP中,快速合成ATP以供给能量。肌酸激酶主要存在于细胞质和线粒体中,与细胞内能量转运、肌肉收缩和ATP再生有着密切联系,能可逆地催化磷酸肌酸和ATP间高能磷酸键的转化。肌酸由L-精氨酸、甘氨酸和甲硫氨酸3种氨基酸合成,L-精氨酸在精氨酸-甘氨酸咪基转移酶(L-arginine glycine amidinotransfercase, AGAT)的作用下断开胍基键形成L-鸟氨酸,切下的咪基转移到甘氨酸上形成GAA,故GAA是一种甘氨酸衍生物。肾脏和胰腺合成的GAA以及外源获得的GAA均通过血液循环运输到肝脏,S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)在胍乙酸-N-甲基转移酶(S-adenosyl methionine: guanidinoacetate N-methyltransferase, GAMT)的催化下将甲基转移到GAA上生成肌酸和S-腺苷同型半胱氨酸(S-adenosyl-L-homocysteine, SAH)[6-8]。SAH提供甲基后水解成同型半胱氨酸和腺苷,其中同型半胱氨酸有2条去路:若甲硫氨酸充足,同型半胱氨酸进入转硫途径,与丝氨酸在胱硫醚β合成酶(Cystathionineβsynthase, CβS)的作用下缩合形成胱硫醚,再水解为L-半胱氨酸和α-酮丁酸;若甲硫氨酸缺乏,同型半胱氨酸从N5-甲基四氢叶酸接受甲基,重新甲基化后生成L-甲硫氨酸(图1)。可见机体SAM的浓度很大程度上决定了同型半胱氨酸和甲硫氨酸的代谢途径。有研究表明,机体内源性合成肌酸所需甲基主要由SAM提供,因此在日粮中补充胍基乙酸势必会增加机体对甲基的需求量[9],然而动物机体不能内源性合成甲基,需要从食物中获得以维持正常的生理功能。甜菜碱作为高效甲基供体,能在甜菜碱-同型半胱氨酸S-甲基转移酶(Betaine-homocysteine S-methyltransferase, BHMT)的催化下,将甲基从甜菜碱转移到同型半胱氨酸上产生二甲基甘氨酸和蛋氨酸,以此满足机体对甲基需求量,同时清除体内同型半胱氨酸的过分积累。LIU等[10]研究发现,在小鼠日粮中添加甜菜碱和胆碱能减少同型半胱氨酸血症的发生率。
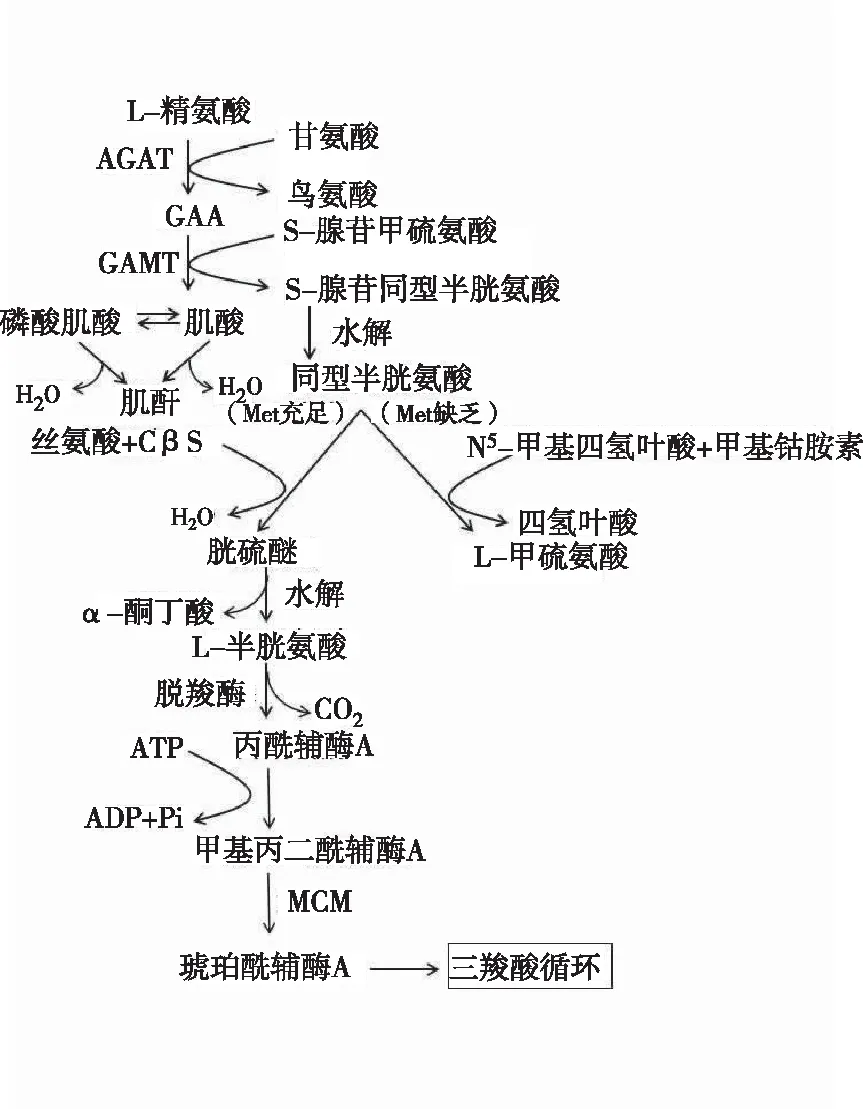
图1 胍基乙酸的代谢途径Fig.1 Metabolic pathways of GAA
2 胍基乙酸在动物生产中的应用
2.1 在家禽生产中的应用
饲粮中外源性添加GAA可直接提高磷酸肌酸ATP值,数值越高说明细胞能量代谢越快,即能为肌肉组织收缩、细胞活动以及合成代谢提供更多及时的ATP[11],从而促进肌纤维的发育,改善动物的生产性能。RINGEL等[12]研究发现,随GAA添加量的增加,肉鸡胸肉的产出量也线性上升。在循环热应激条件下,肉鸡日粮添加GAA(正常蛋白水平的90%)能降低生长肉鸡在高温条件下的死亡率,同时在日粮中添加5 g/kg GAA可以改善热应激条件下肉鸡的生长性能、死亡率、抗氧化状态和肠绒毛形态[13]。张德福等[14]在肉仔鸡基础饲粮中添加800 mg/kg、1 600 mg/kg、4 000 mg/kg和8 000 mg/kg GAA发现,饲粮中添加800~4 000 mg/kg GAA能显著提高肉仔鸡的日增重,降低后期(22~42日龄)和全期料重比;添加800~8 000 mg/kg GAA对肉仔鸡血常规指标、碱性磷酸酶、尿素氮、总胆红素和谷草转氨酶等血清生指标、脏器指数以及组织同型半胱氨酸含量无显著影响,说明肉仔鸡可耐受8 000 mg/kg GAA。
王亚琼等[15]研究GAA对樱桃谷肉鸭抗氧化能力的影响发现,饲粮中添加0.05% GAA的试验组血清中谷胱甘肽过氧化物酶(GSH-Px)活性和还原型谷胱甘肽(GSH)含量均显著增加,肝脏中超氧化物歧化酶(SOD)活性和GSH含量也显著增加,说明在肉鸭饲粮中添加0.05% GAA可在一定程度上提高机体的抗氧化能力。HIRAMATSU[16]研究认为,GAA既可以是抗氧化剂,也可以是助氧化剂。原因一方面是GAA的共轭碱能提供电子而生成自由基-过氧化物,具有促氧化作用;另一方面,GAA代谢后的肌酸和精氨酸有清楚自由基的功能,从而起抗氧化作用。然而目前对 GAA和细胞氧化-抗氧化体系之间的关系还没有明确定论。
2.2 胍基乙酸在家畜生产中的应用
GAA作为肌酸的唯一前体物质,能直接提高磷酸肌酸和ATP等高能量物质的存贮量或利用效率,减少蛋白质和脂肪的分解供能,有效促进肌肉的能量代谢,从而促进动物机体的生长发育[17]。赵元等[18]报道,在育肥猪日粮中添加500 mg/kg、1 000 mg/kg和1 500 mg/kg的GAA能显著提高育肥猪的日增重、胴体重、瘦肉率、屠宰率和眼肌面积,降低料肉比和第十肋骨处背膘厚;与对照组相比,试验组的背最长肌和半腱肌的滴水损失、蒸煮损失、剪切力和粗脂肪含量均显著下降,pH明显升高。刘洋等[19]研究发现,在育肥猪日粮中添加1 g/kg GAA及1 g/kg GAA+0.5 g/kg甜菜碱能显著降低肌肉的滴水损失、硬度和蒸煮损失,同时提高pH45min和pH24h值、肌酸、磷酸肌酸以及ATP含量。这是由于GAA可间接为肌肉的收缩及新陈代谢提供ATP,延缓宰后肌肉糖酵解产生乳酸的速度,即减缓pH的下降,从而提高肌肉系水力改善肉质[20]。
在滩羊基础饲粮中添加0.1%氮氨甲酰谷氨酸(NCG)+0.08% GAA能显著提高试验滩羊的平均日增重、饲料转化率、屠宰率和净肉率,明显改善滩羊体脂的分配,即尾脂和GR值下降,肌内脂肪含量升高[21]。肌酸能够促进肌肉组织蛋白重链和肌动蛋白的合成,加强肌内脂肪组织能量供给,从而提高肌肉中脂肪含量[22]。此外,辛均平等[23]采用体外产气法研究不同水平GAA(0.1%、0.2%、0.3%、0.4%)对锦江黄牛瘤胃体外发酵参数的影响发现,在日粮中添加胍基乙酸能促进瘤胃的发酵功能,试验组瘤胃pH显著低于对照组,瘤胃微生物蛋白含量与GAA添加量呈正相关,随着添加量的增加瘤胃液中挥发性脂肪酸总量显著高于对照组。
2.3 胍基乙酸在水产生产中的应用
有研究发现,在鲤的基础饲料中添加3% EM菌+250 mg/kg GAA可在一定程度上提高鲤的生长性能,促进能量代谢,改善其非特异性免疫力[24]。洑琴等[25]研究GAA对建鲤生产性能和肌肉能量代谢的影响结果表明,在基础饲料中添加250 mg/kg、500 mg/kg和1 000 mg/kg GAA可显著降低饵料系数,其中以添加250 mg/kg GAA能获得较佳的饲料转化率;添加250 mg/kg和500 mg/kg GAA能显著提高肌糖原含量,说明葡萄糖更多以糖原的形式贮存在肌肉中,同时丙酮酸激酶、琥珀酸脱氢酶活性及肌酸激酶活性显著下降,说明外源性添加GAA能直接增加磷酸肌酸和ATP含量,减少葡萄糖分解供能,从而导致糖酵解反应过程的关键酶丙酮酸激酶和三羧酸循环中的标志酶琥珀酸脱氢酶活性降低[26]。
3 展望
GAA作为新型饲料添加剂,不仅能促进动物生产性能、提高饲料利用率、改善肉质,同时还在抗氧化应激方面发挥着一定效果。目前GAA在我国动物生产中的应用尚处于起步阶段,对于其在饲料中的添加剂量及作用机制,特别是对于反刍动物特殊的消化道结构,在日粮中添加GAA在瘤胃内是否会被微生物大量降解,是否遵循单胃动物及家禽同样的代谢机制,或许将是营养学家下一步的研究方向和重点。
猜你喜欢
分子催化(2022年1期)2022-11-02
石油化工(2022年9期)2022-10-19
中老年保健(2022年3期)2022-08-24
中学生数理化·高一版(2022年4期)2022-05-09
中学化学(2021年11期)2021-12-09
昆明医科大学学报(2021年1期)2021-02-07
祝您健康(2020年5期)2020-05-14
健康博览(2020年2期)2020-02-27
食品界(2016年10期)2016-09-10
食品工业科技(2014年6期)2014-05-10
