
宿主整合因子对伤寒沙门菌动力的影响
2021-03-12 07:46:52徐国新朱晓珏王芳陈龙
江苏大学学报(医学版) 2021年1期
徐国新,朱晓珏,王芳,陈龙
(1. 苏州大学附属张家港医院检验科,江苏 张家港 215600; 2. 张家港市中医医院检验科,江苏 张家港 215600)
伤寒沙门菌(Salmonellaentericaserovar Typhi,S. Typhi)属于肠杆菌科沙门菌属,常通过污染的食物或水进入人体消化道,进而引起全身性的感染症状——伤寒热[1-2]。伤寒沙门菌经口摄入,经胃酸、小肠高渗透压等不利环境因素后侵入回肠、盲肠及结肠末端。鞭毛是细菌的动力装置,在伤寒沙门菌致病过程中起着重要作用。鞭毛帮助细菌抵抗环境因素杀伤的同时,参与伤寒沙门菌对肠上皮细胞的侵袭[3]。伤寒沙门菌的鞭毛调控网络复杂,受多种全局调控因子甚至非编码RNA调控[4-6]。宿主整合因子(integration host factor,IHF)作为伤寒沙门菌的全局调控因子之一,含有HimA、HimD两个亚基。IHF能调控伤寒沙门菌多种基因尤其是毒力基因的转录表达[7]。但是,IHF对于伤寒沙门菌动力的影响及鞭毛基因的调控作用仍不清楚。本研究通过分析IHF对鞭毛基因的调控作用,初步探讨其对伤寒沙门菌动力的影响。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 伤寒沙门菌野生株(GIFU10007)、缺失株△himA、△himD为本实验室保存;pBAD33质粒由江苏大学黄新祥教授惠赠;异源表达载体pBAD33用以构建载体himA-pBAD、himD-pBAD,分别转化入△himA、△himD构建相应回补株himA-C、himD-C;空载体pBAD33转化入野生株及缺失株作为空白对照。
1.1.2 主要试剂 限制性内切酶XhoⅠ、EcoR Ⅰ及T4DNA连接酶、反转录试剂盒、DL2000 DNA标准参照物、反转录酶及嵌合荧光染料购自TaKaRa(大连)公司;高保真DNA聚合酶、TaqMix购自南京诺唯赞公司;质粒提取试剂盒、胶回收试剂盒(美国Axygen公司);氯霉素(美国Sigma公司);PCR产物纯化试剂盒(德国Qiagen公司);Trizol试剂及RNA纯化试剂盒(美国Invitrogen公司)。
1.1.3 主要仪器 凝胶成像系统(美国Gene公司,型号Gene Genius Bio Imaging System);PCR仪(美国ABI公司,型号ABI 2700);电转化仪(美国Bio-rad公司,型号GENE PULSER Ⅱ);荧光定量PCR仪(美国Bio-rad公司);微量核酸分光光度计(美国Thermo公司,型号NanoDrop-1000);引物由苏州泓迅生物公司合成。
1.2 方法
1.2.1 回补株构建 以S. Typhi GIFU10007野生株作为模板,使用上游引物:5′-CGCTCGAGATGGCGCTTACAAAAGCTGA-3′(下划线为XhoⅠ酶切位点)与下游引物:5′-CGGAATTCTTACTCTTCTTTGGGCGA-3′(下划线为EcoR Ⅰ酶切位点)PCR扩增得到himA的编码区域序列,使用上游引物:5′-CGCTCGAGATGACCAAGTCAGAATTGAT-3′(下划线为XhoⅠ酶切位点)与下游引物:5′-CGGAATTCTTAACCGTAAATATTGGCGCGAT-3′(下划线为EcoRⅠ酶切位点)PCR扩增得到himD的编码区域序列;PCR产物经纯化后与异源表达载体pBAD33分别酶切,酶切产物经T4连接酶连接成重组质粒,PCR筛选阳性载体,测序验证正确即成功构建回补质粒;回补质粒电转化导入伤寒沙门菌野生株,PCR验证携带了重组质粒的克隆,即构建成功回补株himA-C、himD-C。野生株导入pBAD33空质粒作为对照,记为WT-pBAD。
1.2.2 动力实验 接种WT-pBAD、△himA-pBAD、△himD-pBAD、himA-C、himD-C单克隆至2 mL LB肉汤,37 ℃,250 r/min振摇培养过夜;次日以1∶100转接种于新鲜LB肉汤,250 r/min振摇培养约2 h至D(600 nm)≈0.4;加入阿拉伯糖(终浓度1 mmol/L)继续培养0.5 h以诱导异源表达载体。取菌液1 μL接种于0.3%琼脂半固体培养基(含1 mmol/L阿拉伯糖)孵育12 h;量取动力圈直径,进行统计学分析;通过动力圈直径比较细菌动力。
1.2.3 细菌总RNA提取及反转录 挑取单菌落WT-pBAD、△himA-pBAD、△himD-pBAD,接种至2 mL LB肉汤,37 ℃,250 r/min振摇培养过夜;以1∶100转接于新鲜LB肉汤,250 r/min振摇培养至D(600 nm)≈0.4;加入阿拉伯糖(终浓度1 mmol/L)诱导0.5 h;4 ℃,4 000 r/min,离心10 min收集细菌。用Trizol提取细菌总RNA,并用RNA纯化试剂盒消化残存的DNA,核酸分析仪分析提取的RNA浓度及纯度;取4 μg RNA,根据反转录试剂盒说明书,将RNA反转录成cDNA,反应体系10 μL,反应条件:25 ℃、15 min,85 ℃、5 s。
1.2.4 实时荧光定量PCR(qRT-PCR)分析鞭毛基因flhD、fliA和fljB反应体系20 μL,其中嵌合荧光染料预混液10 μL、上下游引物各0.8 μL、cDNA模版2 μL,双蒸水6.4 μL,以上反应液配置均在冰上进行。反应条件:95 ℃预变性30 s;95 ℃ 5 s,60 ℃ 20 s,共40个循环。利用2-△△Ct计算法得到mRNA相对表达量。所用引物序列如下:flhD上游引物5′-AGCGTTTGATCGTCCAG-3′,下游引物CGTCCACTTCATTGAGCA-3′;fliA上游引物5′-CGACCGATATGACGCTTTGC-3′,下游引物5′-TTCCCGCCACTCATCGTAAG-3′;fljB上游引物5′-CAACCGCTAGTGATTTAGTTT-3′,下游引物5′-CTGTCCCTGTAGTAGCCGTAC-3′;5S上游引物5′-TTGTCTGGCGGCAGTAGC-3′,下游引物5′-TTTGATGCCTGGCAGTTC-3′。每组实验至少重复3次。
1.3 统计学分析

2 结果
2.1 伤寒沙门菌回补株himA-C和himD-C的构建
结果如图1所示,成功构建含有himA、himD编码片段的pBAD33回补质粒,并经测序验证正确。将回补质粒利用电转化的方式导入相应基因缺失株,即成功构建回补株himA-C、himD-C。
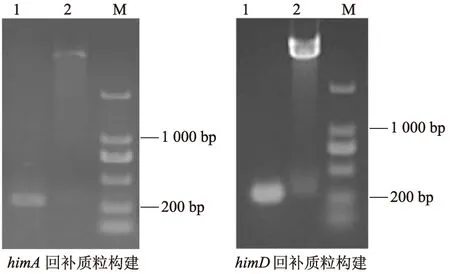
1:PCR验证;2:质粒酶切验证;M:DNA标准参照物
2.2 himA和himD缺失后对伤寒沙门菌动力的影响
动力实验结果如图2显示,与WT-pBAD相比,△himA-pBAD与△himD-pBAD的动力圈直径明显减小(t=14.00,10.58,P均<0.05),himA-C、himD-C的动力圈与WT-pBAD基本一致,差异无统计学意义(P>0.05)。结果表明,himA和himD对伤寒沙门菌的动力起促进作用。
2.3 himA和himD对伤寒沙门菌鞭毛基因mRNA表达的影响
qRT-PCR结果如图3显示,△himA-pBAD中鞭毛基因flhD、fliA、fljB的mRNA表达水平均明显低于WT-pBAD(t=9.55,8.1,11.25,P<0.05或<0.01);△himD-pBAD中flhD、fliA、fljB的mRNA表达水平同样明显低于WT-pBAD(t=10.2,8.1,12.5,P<0.05或<0.01)。由此表明,himA和himD均能够正向调节伤寒沙门菌鞭毛基因flhD,fliA和fljB等mRNA表达。

图2 伤寒沙门菌动力实验

图3 qRT-PCR检测各菌株中鞭毛基因flhD,fliA和fljB的mRNA表达
3 讨论
IHF作为全局调控因子,在多种细菌的基因调控网络中起重要重用,参与致病进程。在新月柄杆菌中,IHF抑制细菌动力;在大肠埃希菌和鼠伤寒沙门菌中,IHF激活细菌动力[8-9]。伤寒沙门菌是一种具有双相鞭毛革兰阴性菌,其鞭毛的构成及功能有特殊性[10]。本研究发现,缺失himA或himD后,伤寒沙门菌动力明显减弱,表明IHF能够促进伤寒沙门菌的动力。
鞭毛是细菌重要的动力器官,在致病过程中发挥重要功能,如细菌定植和侵袭、感染位点的维持、感染后的扩散等[11]。伤寒沙门菌鞭毛的调控机制复杂,有50余个基因参与鞭毛的合成,按照转录调控级别可分为三级。本研究选取的flhD、fliA和fljB分别属于伤寒沙门菌的一、二、三级鞭毛基因,其中fljB为伤寒沙门菌GIFU10007所特有H:z66的编码基因[10]。本研究结果显示,himA和himD均能够正向调节伤寒沙门菌鞭毛基因flhD,fliA和fljB等mRNA表达。
调控因子对靶基因的调节方式有直接与间接两种方式,前者是指调控因子能够直接结合到靶基因的启动子序列,调节转录;后者则通过调节其他调控因子,影响靶基因转录[12]。后续研究中,我们将表达IHF蛋白,分析IHF对于鞭毛基因的调控方式。
综上所述,本研究初步揭示宿主整合因子IHF能够增强伤寒沙门菌的动力,并正向调节鞭毛基因的转录。
猜你喜欢
中国饲料(2022年5期)2022-04-26 13:42:36
中国动物传染病学报(2021年3期)2021-07-21 03:19:30
实用检验医师杂志(2019年3期)2019-11-01 04:21:48
人人健康(2019年10期)2019-10-14 03:25:12
首都公共卫生(2018年5期)2018-11-21 07:02:02
食品科学(2018年10期)2018-05-23 01:27:28
科学家(2017年20期)2017-11-10 23:10:19
科技资讯(2016年19期)2016-11-15 10:39:37
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
