
天皇海山典型分布经济鱼种研究进展
2021-03-09 05:26杨嘉樑李灵智屈泰春黄洪亮
渔业信息与战略 2021年1期
杨嘉樑,李灵智,饶 欣,刘 健,屈泰春,黄洪亮
(中国水产科学研究院东海水产研究所,农业农村部远洋与极地渔业创新重点实验室,上海 200090)
深远海因其丰富的自然资源日益成为各海洋国家和地区进行博弈的重要场所[1-2],而海山作为深远海里特殊的海底生境也因此受到越来越多的关注。海山一般是指在海底隆起至海面以下,高度在1 000 m以上的海底高地[3],一般是由于火山爆发或地壳板块运动而形成[4-5]。由于海山的特殊地质构造影响着海山海域内沉积物的沉积速率、营养物质的丰度和浮游动物及鱼类的栖息模式[6],因此,在海山栖息的生物,诸如鱼类,与其他海域栖息的鱼类有显著差异,研究海山区域的重要鱼类对了解海山这一特殊生境对生物的聚集机制具有重要意义。天皇海山海域作为曾经产量较高的渔场,其海域内栖息的主要经济鱼种的渔业资源丰度与海山这一特殊环境的关系也一直是许多研究关注的热点。
1 天皇海山概况
天皇海山位于西北太平洋的东部,从阿留申群岛一直延伸至夏威夷群岛西北部的海山群,在天皇海山的主要海山中,由北向南以日本历代天皇年号称呼的主要海山有明治、天智、神武、推古、昭和、用明、仁德、应神、光孝、钦明、雄略和恒武等海山,此外,还有科拉汉和汉考克海山,整个海山水域跨度达1 700 n mile(图1)[7-12]。天皇海山的海底深度通常为240~2 800 m,但其中仅有山顶平坦、且山顶至海平面距离在300~500 m的海山可以作为底拖网渔场,因此日本把这些具有宽广平顶的海山称为“渔海山”[7-8]。在海山的斜坡海域及水深较深的水域则不适宜采用底拖网进行捕捞,取而代之采用底刺网进行作业[9]。大洋拟五棘鲷(Pseudopentaceros wheeleri)及红金眼鲷(Beryx splendens)是天皇海山底层水域栖息的典型代表鱼种并成为底拖网及底层刺网的主要捕捞目标[10-11]。
2 大洋拟五棘鲷相关研究进展
2.1 分类
栖息在天皇海山海域资源量最多的鱼种为大洋拟五棘鲷,属鲈形目(Perciformes)鱼类[13],起初有研究[14-15]认为大洋拟五棘鲷与另一种栖息于南半球且形态相似的Pentaceros richardsoni为同一种鱼,但是HUMPHREYS[16]的研究提出,大洋拟五棘鲷与P.richardosoni并非为同一种鱼类,因此重新命名为Pseudopentaceroswheeleri。同一研究还发现,在天皇海山海域所见到的大洋拟五棘鲷有2种体型:一种体高较高,一种体高较矮(图2),通过同工酶性状对两种体型的大洋拟五棘鲷进行分析,结果显示,这2种体型的大洋拟五棘鲷之间没有显示出遗传差异,因此属同一种鱼。
2.2 食性
在表层栖息的大洋拟五棘鲷幼鱼主要以桡足类浮游动物为主要饵料生物,偶尔以翼足类等其他浮游动物为食[14]。进入成鱼期后,浮游阶段的大洋拟五棘鲷的主要饵料生物是磷体虫(原索动物)、纽鳃樽(原索动物)和塔形长角螺等软体动物,此外,褶胸鱼科等小型鱼类、磷虾类等甲壳动物也是大洋拟五棘鲷的捕食对象[17]。但进入海底栖息阶段后,大多数栖息在海底的大洋拟五棘鲷很少进食,因此在海底捕获的大洋拟五棘鲷的胃中鲜有胃含物[17]。但也有其他研究发现,海底栖息的大洋拟五棘鲷通常以甲壳类、翼足类、海蛰、沙丁鱼甚至海藻等为食[14,18-20]。
2.3 繁殖
雌性大洋拟五棘鲷一般达到3龄后才进入产卵期,佐佐木[18]从每月雌性大洋拟五棘鲷生殖指数(gonadosomatic index,GSI)的变化及对其生殖腺的观察中发现,大洋拟五棘鲷的产卵季节应为每年的冬季。 YANAGIMOTO 和HUMPHREYS[21]则通过对生殖腺组织的观察,提出大洋拟五棘鲷的产卵月份为当年11月至次年2月。
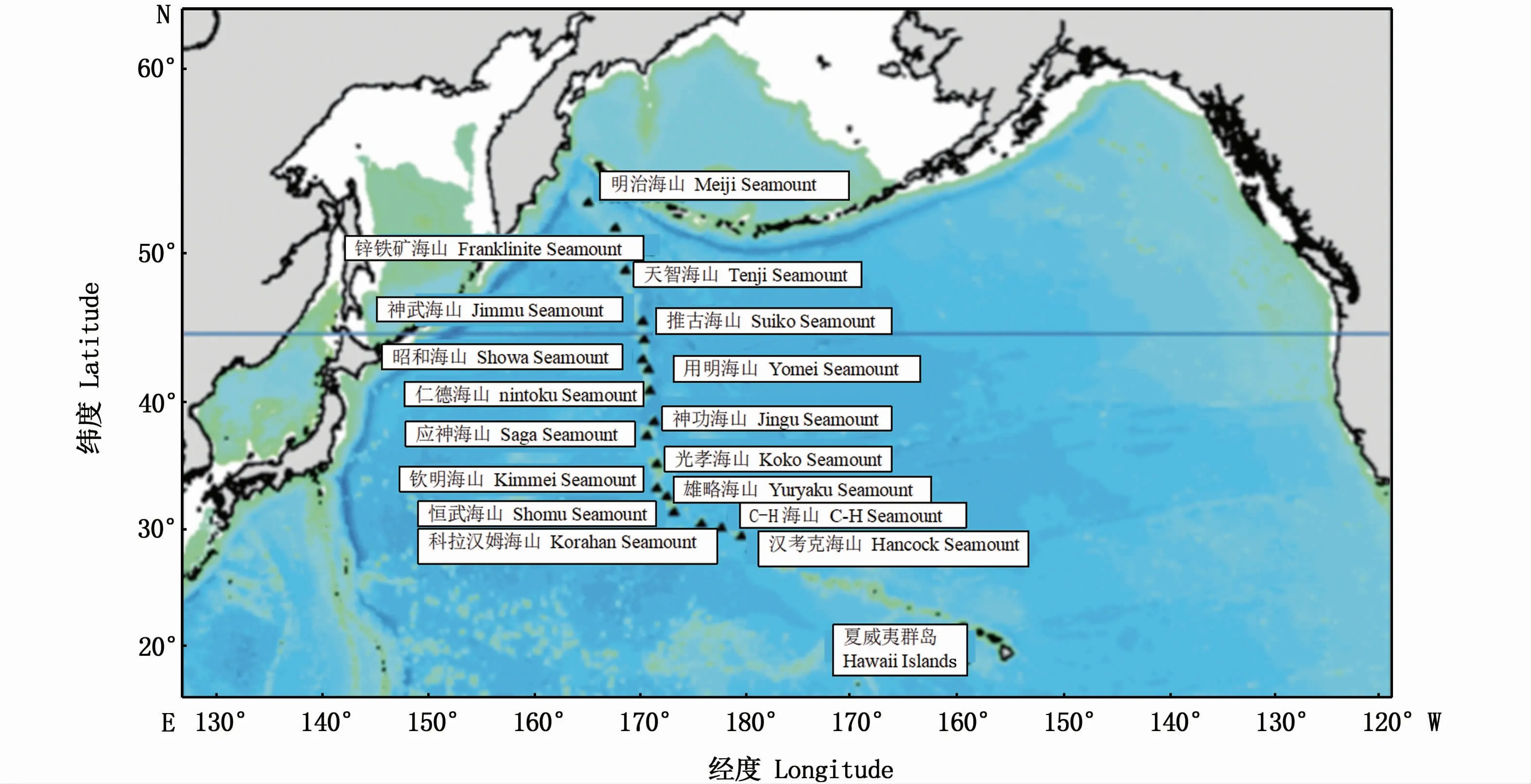
图1 天皇海山海域主要海山群[12]Fig.1 M ajor seamounts in Em peror Seamount waters

图2 大洋拟五棘鲷Fig.2 Pseudopentaceros wheeleri
2.4 种群分布特征
天皇海山海域栖息大洋拟五棘鲷成鱼,主要分布在水深300~500m处的海山海域。然而,根据相关调查,发现在北美洲加利福尼亚半岛以西海域、日本本州中部南岸以南的伊豆群岛、小笠原群岛、九州以南海域及帕劳群岛等周边的海山海域也分布有大洋拟五棘鲷的少数个体[13](图3)。大洋拟五棘鲷于每年11月至次年2月在天皇海山海域内产卵;1—3月,鱼卵及孵化的仔稚鱼分布于天皇海山的表层水域[22-23];进入幼鱼阶段后,大洋拟五棘鲷游动至40°~50°N、140°~180°W 附近的表层海域,维持约2年的浮游生活[16,24-26]。在浮游期终止后,大洋拟五棘鲷随即下沉至海山的海底区域并进入底栖阶段栖息,从附着于大洋拟五棘鲷鳃部的寄生虫状况看,成鱼进入海底栖息的月份可能为5—7月[27]。
通过PCR-RFLP分析天皇海山海域内各海山采集的大洋拟五棘鲷样本的mtDNA发现,栖息在各海山的大洋拟五棘鲷群体之间无遗传性差异[28]。柳本和北村[29]对从天皇海山海域、太平洋表层海域和八丈岛(伊豆群岛的南侧)海域采集来的大洋拟五棘鲷样本进行了mtDNA的PCR-RFLP分析,也未检出遗传性差异。由此可以确认在天皇海山各海域内栖息的大洋拟五棘鲷之间没有遗传性差异,属于同一种群[30]。如前所述,虽然冬季在各海山出生的仔稚鱼在次年3月仍栖息在海山上的表层海域,但当其进入到幼鱼期时便广泛地分布于北太平洋,并于2~3年以后返回到天皇海山海域[25]。
2.5 生长与死亡特征
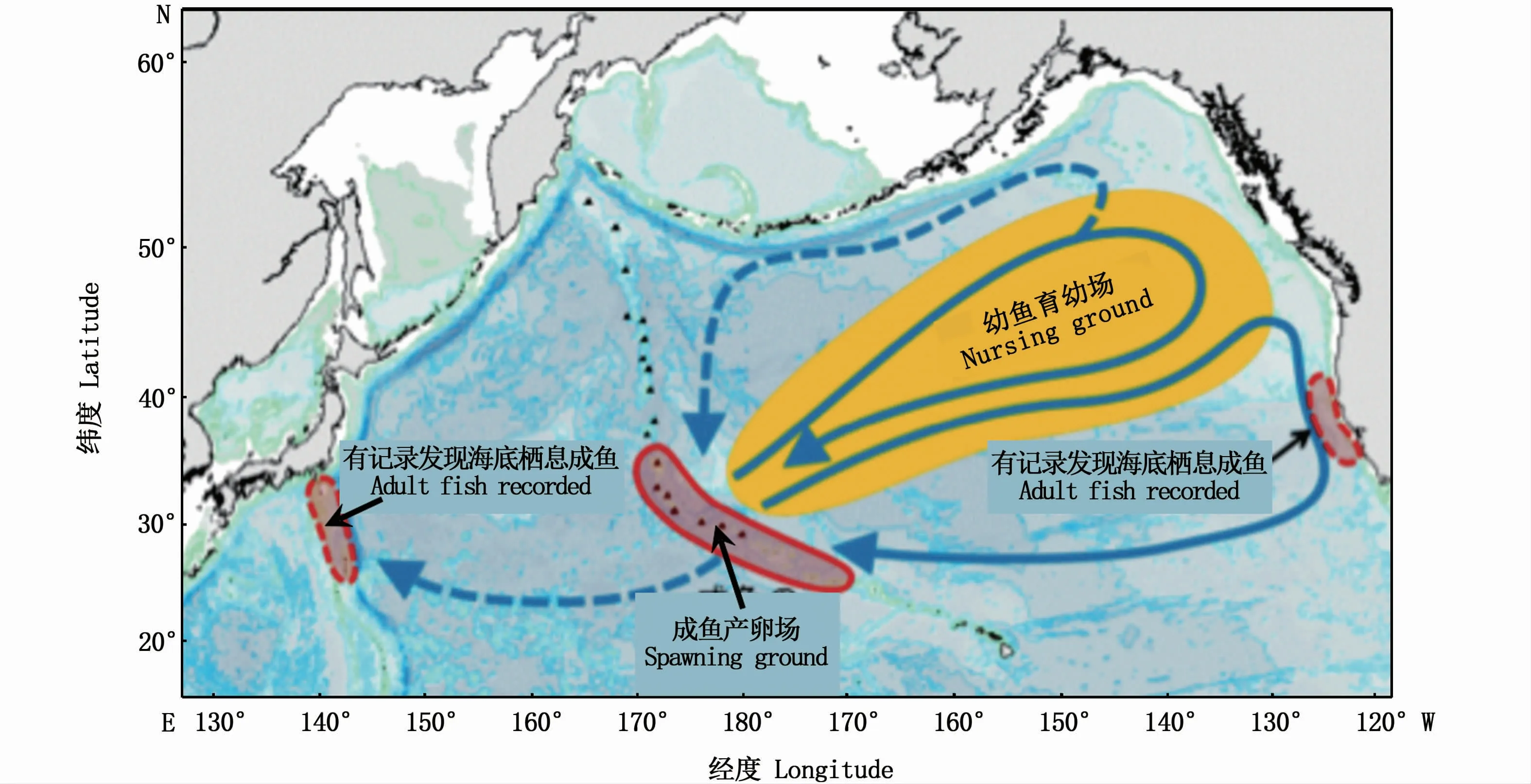
图3 大洋拟五棘鲷的产卵场和成育场Fig.3 Spawning ground and nursing ground of PseudopentacerosWheeleri
大洋拟五棘鲷在浮游阶段,其体长与年龄具有一定相关性,比如未满周岁的鱼苗其体长一般为4~7 cm,1龄鱼的体长则一般为18~21 cm,2龄鱼的体长则一般为25~28 cm。在天皇海山海域表层栖息的大洋拟五棘鲷其体长一般约为30 cm,体质量约为600 g。当大洋拟五棘鲷进入海底栖息阶段后,其沿体轴方向的生长几乎停滞,这些表层浮游群体的年龄约为2.5~3.5龄[31],因此,底栖的大洋拟五棘鲷并不能通过体长组成推定年龄并区分年龄组。由于大洋拟五棘鲷的幼鱼样本可以通过耳石切片的日轮推定其孵化日,因此也可以通过耳石切片的周日推断底栖阶段成鱼的年龄。
此外,在海山着底后的大洋拟五棘鲷,为了产卵而将其在浮游阶段贮存在体内的脂肪逐步消耗,导致其体型逐渐消瘦,致使其体高及体宽等参数出现逆生长现象,从而发生如图2所见浮游群体与底栖群体在形态上的显著差异。SOMERTON和KIKKAWA[32]用体长和体高的比值(fatness index,FI),即FI指数的长年变化对年龄群的动向进行推测,认为如某一年体高较高的个体(即FI值高的个体)数量较多的话,则认为该年大洋拟五棘鲷资源补充量较多。之后根据调查FI值逐年变小的过程,进而推测当年补充群体的寿命。根据推算结果,大洋拟五棘鲷的自然死亡系数(natural mortality coefficient)M 约为0.54,这是一个非常大的值,大洋拟五棘鲷在海山着底后,经推测要5年左右才会死亡,因此,大洋拟五棘鲷寿命推算为7~8年[16]。
3 红金眼鲷相关研究进展
3.1 分类
红金眼鲷(图4),为金眼鲷科(Berycidae)鱼类,原先认为其与另一种金眼鲷科的同属鱼类软体金眼鲷(B.mollis)为同一种[33],但后经形态学及遗传学等研究发现,两者之间存在明显的差异[34],因此后续对两种鱼类加以区分,如通过背部软质鳍条数量、呼吸孔附近鳞片形状及排泄孔位置加以区别[35]。
3.2 食性
成年红金眼鲷一般以灯笼鱼科(Myctophidae)鱼类、头足类及甲壳类等浮游动物为饵料生物[36],有研究认为体长越长的红金眼鲷其胃含物中鱼类的占比越高[37]。红金眼鲷幼鱼的食性目前仍然尚未明确。对此前在天皇海山捕获的红金眼鲷样本进行解剖,发现其空腹率仅为10%,其胃含物主要包括磷虾、翼足类、小型头足类及海藻等[38]。

图4 红金眼鲷Fig.4 Beryx splendens
3.3 繁殖
红金眼鲷繁殖群体的年龄组一般为4~5龄。天皇海山海域初次进入繁殖期的红金眼鲷,其体长一般为28~35 cm[39],处于繁殖期的红金眼鲷的优势体长组则为31~35 cm[40]。另外,在不同海域繁殖群体中的优势体长组的范围有所差异,例如在智利沿海海域的红金眼鲷其处于繁殖期的群体优势体长组为40~44 cm[41]。红金眼鲷在其繁殖期内可进行多次产卵,具体产卵时间也因不同的海域有所差异。由于目前天皇海山区域进行的资源调查中一直未发现红金眼鲷的鱼卵样本,因此相关研究根据所采集到的红金眼鲷仔稚鱼及幼鱼样本的耳石日轮及生殖腺指数等推算出天皇海山海域栖息的红金眼鲷产卵期应该为每年的7月[42]。
3.4 种群分布特征
相关研究根据各海域采集到的红金眼鲷线粒体DNA的部分核苷酸序列进行分析,发现栖息在大西洋、印度洋太平洋间及北大西洋的红金眼鲷种群之间已经出现了明显的遗传分化现象,然而栖息在印度洋及太平洋的红金眼鲷种群间却未发现这种遗传分化[43-44]。日本学者通过标志放流及微卫星技术对栖息在日本附近海域的红金眼鲷进行跟踪[45],发现红金眼鲷可能会随黑潮的流动方向出现远距离且大规模的洄游现象,因此在日本近海所捕获的红金眼鲷均属同一种群,相关遗传学研究也同样证实日本近海所捕获的红金眼鲷之间没有遗传差异[46]。由于红金眼鲷在孵化后150~300 d为其浮游栖息阶段,且在其进入海底栖阶段后仍具有长距离洄游的特征,因此有部分学者提出假说,认为天皇海山栖息的红金眼鲷与日本近海的红金眼鲷可能也属于同一种群并存在洄游现象[47],但是到目前为止,这种假说尚无任何研究结果佐证。所以,天皇海山海域栖息的红金眼鲷目前仍然被认为是一个独立的种群。
3.5 生长与死亡特征
红金眼鲷的年龄通常是根据耳石切片的日轮进行推算,并据此推算出红金眼鲷年龄与体长的关系,红金眼鲷在1龄时的体长约为16~22 cm,在3龄时其体长增长至24~28 cm,5龄鱼的体长则为28~32 cm,10龄鱼的叉长为37~41cm,红金眼鲷的寿命达到15龄时,其体长为41~46 cm[35]。研究发现,通过耳石鉴定发现的寿命最长的红金眼鲷为26龄[44]。柳本[38]也采用了耳石鉴别的方法对天皇海山海域栖息的红金眼鲷的年龄进行研究,结果显示天皇海山海域栖息的红金眼鲷生长速率和其他海域的红金眼鲷生长速率无显著差异,但其采样样本的年龄范围绝大多数为1~4龄的个体。因此有学者认为,根据这个结果,天皇海山红金眼鲷的生长速率可能被高估,如高桥[42]通过研究更广年龄范围(1~13龄)的红金眼鲷样本后发现,栖息在天皇海山的红金眼鲷幼鱼生长速率相较于栖息在其他海域的红金眼鲷幼鱼明显更为缓慢。
4 天皇海山底栖鱼类聚集机制分析
影响海山区域生物丰度的环境因素众多,其中水深是一个重要指标。天皇海山形成鱼群高密度聚集的海山,其山顶通常位于海平面下的真光层,因此海山山顶区域易成为生物高度聚集的区域。如果海山穿过缺氧区,海山顶部的生物将会非常稀少,但海山的陆坡位置则反而更易成为生物高度聚集的区域[48]。这也是天皇海山部分具备商业开发价值的渔场同时存在使用底层拖网及底层刺网两种作业方式对大洋拟五棘鲷及红金眼鲷进行捕捞的重要原因。
由于维持海山海域高生物量的形成机制无外乎上升流、水平海流输送及地形诱集这三种机制[49]。传统观点认为海山区域的高生产力是由于海山海域上升流和泰勒柱的混合作用导致,其中上升流是指在海流经过隆起的海山地形时将海山底部富含高营养盐的海水输送至海山顶部的过程[49]。当海山顶部位于真光层所在范围时,海水中的浮游植物将利用上升流输送的营养盐进行光合作用,并在泰勒柱的影响下,导致海山初级生产力提高并被限制在海山上方海域,从而吸引海山海域其他海洋生物的聚集。而泰勒柱是指环绕在海山附近的特殊环流,即当稳定的海流在流经海山时,其原本的流速及流向被海山地形破坏而加速流动,同时受地球自转的影响在海山周围形成的孤立环流。
如果海山山顶穿越真光层并处于缺氧区的水层,则上升流几乎不能穿越真光层并在海山顶部区域长期停留,因此对浮游植物生长的影响有限。这也是部分存在上升流机制的海山海域,其山顶部分的叶绿素浓度反而低于周边开阔水域的叶绿素浓度[50-51]的原因。在这种情况下,海山海域内的高生物量极可能是由于海流与海山地形相互作用而导致外来的营养物质、浮游动物及鱼类等生物的输入所引发,即海流水平输送效应,这种水平海流在流经海山时受到泰勒柱机制的作用而在海山周围形成环流并促进海底颗粒物质的再次悬浮,因此这种效应对维持海山附近海域的高生物量同样起着关键作用。
综上所述,在天皇海山栖息的大洋拟五棘鲷及红金眼鲷,其高生物量形成的机制可能因不同的海山而有所差别,如在以底拖网为主要生产方式的海山海域,该海山海域内的大洋拟五棘鲷及红金眼鲷的高生物聚集丰度可能主要受上升流机制控制,而在部分采用底刺网进行捕捞的渔场,其海山生物资源的丰度则可能受海流水平输送机制所控制。
5 结语
海山生态系统规模庞大,因此可以形成具有开发价值的大型渔场,但其渔业资源种群本身也十分脆弱。以天皇海山为例,20世纪60年代末大洋拟五棘鲷的年捕捞量虽然高达17×104t,但随后其资源量迅速下降,至今仍处于衰退阶段[52];红金眼鲷虽然仍具备一定规模的产量,但近年来其产量却已出现连续下滑的现象[12],说明人类的活动已经对天皇海山的重要渔业资源产生了深远影响。由于大洋拟五棘鲷及红金眼鲷是天皇海山栖息鱼类中的主要代表鱼种,其资源状况对保持当地生态系统稳定发挥着重要作用,因此如果这些在海山栖息的鱼类资源出现衰退甚至崩溃,势必对天皇海山整个海洋生态系统造成巨大影响。鉴于海山生态对于全球生态体系的重要作用,而天皇海山亦是具备高生物量聚集的海山系统,因此,对该区域内大洋拟五棘鲷和红金眼鲷等具有高生物聚集特点的经济鱼种的研究将有助于理清这些物种的生活史,弄清海山环境与这些物种分布、扩散成因的耦合关系,并有助于加深对天皇海山这一特殊生境对生物多样性和生态系统影响的整体认知。
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
37°女人(2019年4期)2019-04-20
故事会(2018年12期)2018-06-23
电子技术与软件工程(2016年20期)2016-12-21
大江南北(2016年3期)2016-11-22
河北渔业(2015年10期)2015-10-15
环球时报(2009-09-16)2009-09-16
幸福·悦读(2009年5期)2009-06-10
祝您健康(1982年1期)1982-12-30
