
GPRASP2基因编辑猪胎儿成纤维细胞系的建立
2021-02-26 09:29鲁雅洁王红顺杨海元魏钦俊戴一凡
南京医科大学学报(自然科学版) 2021年1期
张 敏 ,姚 俊,鲁雅洁,王红顺,王 盈,杨海元,魏钦俊,曹 新*,戴一凡
1南京医科大学医学遗传学系,江苏 南京 211166;2南京医科大学江苏省异种移植重点实验室,江苏 南京 211166
G 蛋白偶联受体(G protein⁃coupled receptor,GPCR)是人类基因组中最庞大的膜蛋白家族,它们是一类具有7 次跨膜螺旋结构的超家族[1],主要通过与配体结合并发生内吞后发挥作用,可调控生物体内许多重要的生理过程。G蛋白偶联受体相关分选蛋白(GPCR associated sorting protein,GASP)主要在GPCR内吞后的分选过程发挥重要作用,通过与受体的羧基端相互结合,从而介导GPCR进入降解或再循环途径,从而调控相关信号转导[2]。GPRASP2 为GASP 家族成员(即GASP2),参与GPCR 活性调节,并与肿瘤发生、细胞生长及衰老、生理调节和溶酶体降解等生理过程相关[2]。本课题组在1 个X 连锁隐性遗传耳聋家系中首次定位和发现GPRASP2 基因突变(c.1717_1718GC>AA,p.A573N)与综合征性耳聋(syndromic hearing loss,SHL)的发生相关[3]。为进一步推进对GPRASP2基因突变/功能缺陷致聋的分子病因学的理解,还需要在模型动物水平进行耳聋基因型/表型相关性的研究和GPRASP2功能的探讨。
随着基因编辑技术的发展,构建基因缺陷的动物模型为从病理与病因学上研究遗传性耳聋提供了有效途径。小鼠等啮齿类模型动物的内耳发育、遗传和解剖结构与人类差别较大[4],无法精确模拟人内耳的生理功能和病理过程。猪在分子进化上与人亲缘关系相近,在遗传、生理生化、器官发育和病程发展等方面,特别在内耳发育、听器结构和功能上与人相似[5-6]:猪出生时耳蜗形态已基本发育成熟并已具备正常听力,这与人胚胎时期的内耳发育过程一致[7-8];猪的内耳耳蜗形态以及螺旋器结构和功能与人相似;猪的听阈和敏感听力范围与人接近[9],具有相似的听性脑干反应(auditory brainstem response,ABR)[10];巴马小型猪体型大小合适,遗传背景明晰,应用于遗传性耳聋的研究更具优势[10]。本研究采用CRISPR/Cas9 介导的基因编辑技术成功获得猪GPRASP2 基因敲除的猪胎儿成纤维细胞(porcine fetal fibroblasts,PFF),为构建GPRASP2 基因缺陷的巴马小型猪模型和在体水平探索GPRASP2 的致聋机制奠定了实验基础。
1 材料和方法
1.1 材料
PFF由南京医科大学江苏省异种移植重点实验室提供;pX330 质粒(Addgene 423230)、带G418 抗性标记的pCMV td⁃tomato 质粒(Clontech 公司,日本);BbsⅠ限制性内切酶和T7EN1 酶(New England Biolabs公司,美国);琼脂糖凝胶回收试剂盒(Qiagen公司,德国);质粒小提中量试剂盒和DH5α感受态细胞(北京天根生化科技公司);Basic NucleofectorTMKits 和细胞转染仪(Lonza 公司,德国);DMEM 培养基、胎牛血清、胰酶、青/链霉素双抗和PBS 缓冲液(Gibco公司,美国);pMD18⁃T载体(TaKaRa公司,日本);引物和磷酸化的寡核苷酸序列由南京金斯瑞公司合成。
1.2 方法
1.2.1 人/猪GPRASP2的生物信息学分析
采用MEGA7.0 软件,选取包括猪和人等20 种物种进行GPRASP2 基因亲缘性分析,并绘制GPRASP2的系统进化树。采用GeneDoc 软件,比对人/猪GPRASP2 氨基酸序列,并进行同源性和保守性分析。
采用DNAstar软件中Protean模块对人/猪GPRASP2 蛋白进行二级结构分析,以Chou⁃Fasman算法预测蛋白α螺旋、β折叠、β转角的比例。采用Swiss⁃Model在线工具对人/猪GPRASP2蛋白的三维结构进行模拟(https://www.swissmodel.expasy.org/),因Protein Data Bank(PDB)数据库中未检索到GPRASP2 同源蛋白的结构,故采用从头计算法对人/猪GPRASP2蛋白主要功能结构域Arm2(含250个氨基酸残基)进行三维结构模拟。
1.2.2 CRISPR/Cas9的靶点设计和打靶载体构建
利用CRISPR 靶点在线设计工具(http://crispr.mit.edu/),分别在猪GPRASP2 基因编码区(coding sequence,CDS)的上游和下游各设计1对20 bp左右的sgRNA 寡核苷酸序列(sgRNA oligo,表1),由南京金斯瑞合成5′端磷酸化修饰的sgRNA oligo。

表1 GPRASP2靶点位置及sgRNA寡核苷酸序列Table 1 GPRASP2 targeting loci and sgRNA oligonucleo⁃tide sequence
载体构建步骤主要包括oligo退火、载体酶切和连接[11-13]。将合成的sgRNA oligo 以去离子水稀释至100 μmol/L,反应体系(10 μL):正链oligo 1 μL,反链oligo 1 μL,去离子水8 μL;PCR 仪设置退火程序:37 ℃孵育30 min,95 ℃孵育5 min,再以5 ℃/min速度逐渐降温至25 ℃;退火后将oligo 稀释250 倍。用BbsⅠ限制性内切酶对pX330质粒进行线性化,并连接退火产物,再转化感受态DH5α,挑单克隆菌落至摇管,37 ℃培养12~16 h,分装1 mL 的菌液送测序。测序验证后冻存菌液,并小提质粒。
1.2.3 细胞转染和单克隆细胞的筛选
PFF培养于含16%胎牛血清的DMEM培养基含中,待细胞汇合度达到90%后,以0.05%胰蛋白酶消化并离心收集细胞;取1 μg pCMV td⁃tomato 分别与空载pX330质粒、靶向猪GPRASP2基因CDS上游和下游的重组pX330⁃CDS⁃UP/pX330⁃CDS⁃DOWN质粒各5 μg 共转染PFF;参照Lonza 核转试剂盒说明书配制核转液,使用程序U⁃023进行核转,转染后24 h加1 mg/mL 的G418 进行药筛培养,隔天换液,并根据细胞生长状态调整G418用药浓度。9~10 d后,显微镜观察单克隆位置,并在皿底标记;弃皿中培养液,PBS清洗2遍,将克隆环放置于皿底单克隆位置,加0.05%胰蛋白酶消化细胞,转接于24孔板中,待细胞长满后再转接至12孔板中培养,孔板盖上标记克隆编号和日期;12 孔板中细胞长满后,取部分细胞用NP40裂解液裂解,提取基因组DNA,PCR 扩增靶点区域,CDS1 上游引物5′⁃TTCTGCACTCTGTTG⁃GCTGAG ⁃ 3′,下 游 引 物5′ ⁃ AGCAGCAGAAC⁃CAGACTCATT⁃3′,扩增产物长度722 bp。反应条件:95 ℃预变性3 min;95 ℃15 s,60 ℃15 s,72 ℃1 min,35 个循环;PCR 产物经切胶回收,连接pMD18⁃T 载体,连接产物转化感受态DH5α中,每板挑取12个菌落送公司测序;测序结果与猪GPRASP2基因序列(XM⁃003135261.4)进行比对,鉴定得每个克隆细胞的基因型。
1.2.4 重组载体打靶效率验证
提取转染细胞的基因组DNA,同前PCR扩增;切胶回收PCR 产物(5 μL),加入2 μL 10×NEBuffer 2.0,用Nuclease⁃free 水补足至19 μL,在PCR 仪中进行退火反应:95 ℃10 min,再逐渐冷却至室温;退火结束后,取9.5 μL 退火产物加入0.5 μL T7EN1酶,剩余9.5 μL 退火产物加0.5 μL 去离子水(即不加酶的对照组),混匀后37 ℃下反应15 min;加入0.5 μL 蛋白酶K使T7EN1酶失活,采用1.5%琼脂糖凝胶电泳分析和检测打靶效率。根据条带灰度值,计算细胞转染敲除效率,敲除效率=100×[1-(1-裂解产物占比)1/2]。
1.2.5 Western blot验证
分别取野生型和GPRASP2基因敲除的PFF,加入含有蛋白酶抑制剂的RIPA 裂解液后超声破碎和低温离心,提取总蛋白,经SDS⁃PAGE 胶分离,转膜至PVDF 膜上,用5%脱脂奶粉室温封闭2 h,加GPRASP2 抗体(1∶1 000,Abcam 公司,英国)和β⁃tu⁃bulin 抗体(1∶1 000,北京翼飞雪),4 ℃孵育过夜,TBST 洗膜后室温孵育二抗(羊抗兔或羊抗鼠)2 h,TBST洗膜后,化学发光仪(Bio⁃Rad公司,美国)检测蛋白表达情况。
2 结果
2.1 人/猪GPRASP2基因的分子进化关系与同源性分析
包含猪和人在内20个物种的GPRASP2系统进化树如图1A 所示,分值越高的节点其置信度也越高。进化树显示人/猪GPRASP2在分子进化上亲缘关系更近,且人/猪GPRASP2蛋白同源性较高,氨基酸序列一致性达86%(图1B),且主要功能结构域Arm2的氨基酸序列一致性高达94%。
2.2 人/猪GPRASP2蛋白结构分析
采用DNAstar 软件中Protean 模块的Chou⁃Fas⁃man 算法对人/猪GPRASP2 蛋白进行二级结构分析。人/猪GPRASP2 蛋白及其主要功能结构域Arm2 的二级结构具有相近的α螺旋、β折叠、β转角的数量(表2),提示人/猪GPRASP2蛋白的二级结构相似。蛋白三维结构模拟显示人/猪GPRASP2的主要功能结构域Arm2的三维结构相似。考虑到人/猪GPRASP2在分子进化上亲缘关系相近且高度同源,推测人/猪GPRASP2 蛋白具有相似的生物学功能,这还需从在体水平进一步验证。
2.3 重组打靶质粒和GPRASP2 基因敲除PFF 的基因型鉴定
测序结果如图2 所示,pX330 中分别插入了GPRASP2基因CDS上游和下游靶点的sgRNA序列。
经测序鉴定,共获得42个阳性单克隆,其中5个为双等位突变的阳性单克隆(表3),包含5种等位基因突变型(图3),4 种导致GPRASP2 编码氨基酸的移码和蛋白截短。
2.4 GPRASP2基因敲除效率的验证
如图4所示,转染pX330⁃CDS⁃UP和pX330⁃CDS⁃DOWN 重组打靶质粒的PFF 对应的PCR 产物均可观察到明显剪切条带,敲除效率分别为20%和15%,选取敲除效率较高的pX330⁃CDS⁃UP重组质粒用于后续转染和单克隆PFF筛选。
2.5 GPRASP2基因敲除PFF的Western blot验证

表2 GPRASP2蛋白及其Arm2结构域的二级结构分析Table 2 Analysis of the secondary structures of GPRASP2 protein and Arm2 domain(%)
采用Western blot 随机验证其中1 株GPRASP2基因敲除PFF细胞系(GPRASP2c.113_114delCT/c.113_114delCT)中GPRASP2 蛋白的表达情况。如图5 所示,在GPRASP2 基因敲除PFF 细胞中未检测到GPRASP2蛋白表达,表明GPRASP2 基因敲除PFF 构建成功,可用于后续的体细胞核移植。

图2 重组打靶质粒测序验证图Figure 2 Sequencing of recombinant vectors
3 讨论

图3 GPRASP2基因敲除PFF的等位基因型分析Figure 3 The allelic genotypes of GPRASP2⁃knockout PFF

表3 GPRASP2基因敲除PFF单克隆细胞基因型Table 3 Genotypes of GPRASP2⁃knockout monoclonal PFF
本课题组在中国1个X⁃连锁遗传性SHL大家系中成功定位和克隆了1 个新的致聋基因GPRASP2,该家系患者均为男性且携带GPRASP2突变,表现为重度以上听力损失伴外耳廓畸形、内耳道狭窄/闭锁以及眼睑下垂等表型,并在家系成员中呈现典型的遗传共分离[3]。GPRASP2 基因定位于人染色体Xq22.1,编码区位于第5 外显子,共编码838 个氨基酸,其羧基端含有高度保守的Arm2功能结构域,该结构域主要通过与多个GPCR的羧基末端结构域相互作用,从而特异性靶向GPCR 进入溶酶体降解或再循环途径[14]。本研究提示人/猪GPRASP2在分子进化上亲缘关系相近并高度同源,且人/猪GPRASP2蛋白及其主要功能结构域Arm2在二级结构和三级结构上高度相似,推测GPRASP2 基因在人/猪相应组织器官(包括内耳)中可能具有相似的生物学功能。

图4 猪GPRASP2基因敲除靶点的T7EN1酶切验证Figure 4 T7EN1 cleavage assay of target sites in porcine GPRASP2
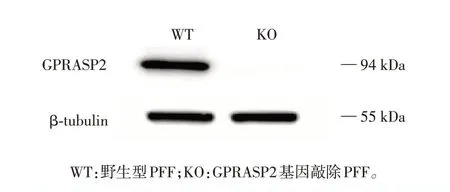
图5 GPRASP2基因敲除PFF的Western blot验证Figure 5 Western blot assay of GPRASP2⁃deficient PFF
GPRASP2作为GASP家族成员(GASP2)在GPCR 内吞后的分选过程中发挥着重要作用。GP⁃CR经配体刺激后,质膜表面的蛋白受体经G蛋白偶联受体激酶磷酸化以及β⁃arrestins 作用下与G 蛋白解偶联,通过网格蛋白有被小泡的协助完成内吞,内吞后的分选途径一般包括再循环和降解[15]。目前已经鉴定出多个受体再循环途径以及GPCR内吞后进入溶酶体降解途径的GASP[15⁃16],涉及多方面的生理功能。有研究表明GASP家族蛋白可作为肿瘤的分子标志物指示肿瘤的发生发展,GASP1在多种类型的肿瘤患者血清中都具有较高的表达量,包括乳腺癌、脑瘤、肝癌以及肺癌等[17];GASP2和GASP3在术前和术后头颈鳞状细胞癌患者中的表达具有显著差异[18];Horn 等[19]研究发现GPRASP2 羧基端部分与亨廷顿蛋白(htt)的氨基端(polyQ)相互作用,形成htt⁃GPRASP2 复合物,参与受体胞吞作用和突触后信号转导,影响受体的转运,而这一过程与亨廷顿舞蹈症发生有较大相关性;Edfawy 等[20]研究发现GPRASP2 能够通过代谢型谷氨酸受体(mGluR)信号通路调控神经系统发育,且敲除GPRASP2基因导致小鼠出现自闭症谱系障碍样行为[20]。现有研究提示GPRASP2与神经系统疾病相关;本课题组则在人类家系中首次关联了GPRASP2基因突变/功能缺陷与SHL的发生相关[3]。
为精确模拟GPRASP2突变导致的SHL表型,从在体水平研究GPRASP2 的功能及其突变致聋的机制,本研究拟选择巴马小型猪构建耳聋动物模型。猪是除了灵长类动物以外在进化关系上与人最接近的实验动物,并在诸多器官(包括听觉器官)的解剖结构和发育遗传方面高度相似[13]。采用基因编辑技术构建基因缺陷的耳聋模型猪可精确模拟人遗传性耳聋的疾病表型和病理过程,如杨仕明课题组利用乙基亚硝基脲(ENU)诱变构建了MITF基因敲除小型猪模型,成功复制了人Mondini 畸形病伴听力损失的临床表型[21];本课题组基于CRIS⁃PR/Cas9 介导的基因编辑技术成功构建了OSBPL2基因敲除的巴马小型猪模型,首次精确模拟了人OSBPL2 基因突变导致的渐进性听力损失的临床表型(DFNA67)和内耳听毛细胞的病理过程[13]。
本研究利用CRISPR/Cas9 技术成功构建了GPRASP2基因敲除的PFF细胞系,为后续体细胞核移植和重组胚胎构建提供合适的供体细胞,为构建GPRASP2 基因敲除的巴马小型猪模型奠定了前期基础,以期为进一步探索GPRASP2基因突变致聋机制与基因治疗提供合适的大动物模型。
猜你喜欢
首都食品与医药(2022年19期)2022-11-19
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
湖北农业科学(2022年11期)2022-07-18
江西农业学报(2021年4期)2021-04-20
昆明医科大学学报(2021年2期)2021-03-29
实用肿瘤学杂志(2020年4期)2020-12-08
三农资讯半月报(2020年11期)2020-06-21
吉林农业(2014年9期)2014-09-18
同位素(2014年2期)2014-04-16
