
妊娠期糖尿病对新生子代大鼠膈肌功能的影响
2021-01-31 08:03张瑞丽杨筱青张晓鸽郭华峰朱继红
吉林大学学报(医学版) 2021年1期
张瑞丽, 杨筱青, 张晓鸽, 郭华峰, 朱继红
(1. 河南省郑州市妇幼保健院产科,河南 郑州 450012;2. 郑州大学第一附属医院产科,河南 郑州450000;3. 吉林大学第一医院生殖中心,吉林 长春 130021)
妊娠期糖尿病(gestational diabetes mellitus,GDM)是指妊娠期首次出现糖耐量受损或糖尿病,是临床上常见的产科疾病[1]。GDM 对母婴均可造成危害,尤其是在妊娠中晚期,母体内分泌系统的变化引起体内各种激素的过多分泌。因此对于GDM 患者,若妊娠期高血糖长期得不到有效控制,母体内的高血糖可以通过胎盘进入胎儿体内,导致胎儿体内也出现高血糖和高胰岛素血症[2]。胎儿体内血糖和胰岛素水平升高,导致胎儿对营养物质需求增加,进而引起胎儿过度或者异常生长,增加母体的剖腹产率和流产率[3]。已有证据[4-5]表明:高血糖和高胰岛素血症可引起胎儿心脏超微结构和功能的改变,或诱导骨骼肌出现胰岛素抵抗。膈肌是一种特殊的骨骼肌,是人体的主要呼吸肌,贡献人体70%~85%的呼吸做功。与成年人不同,新生儿的呼吸模式以小潮气量高呼吸频率为特点[6]。正常的膈肌功能对新生儿的呼吸过程至关重要;反之,膈肌功能异常会导致呼吸功能不全,潜在性地影响新生儿的呼吸功能。而GDM 对新生子代大鼠膈肌功能的影响尚未见报道。本研究通过构建大鼠GDM 模型,观察新生子代大鼠膈肌收缩力、纤维横截面积和超微结构的变化,并检测膈肌组织中氧化应激标志物水平,为研究GDM 对新生子代大鼠膈肌功能的影响提供依据。
1 材料与方法
1.1 实验动物、主要试剂和仪器8 周龄SPF 级SD 大鼠,雌性50 只,雄性25 只,购自郑州大学动物实验中心,动物生产许可证号:SCXK (豫)2017-0001。饲养于温度约25 ℃、湿度约50%、12 h 昼夜交替的动物房里,自由进食水。链脲佐菌素(STZ) 购自美国Sigma 公司,一抗(MY-32、NOQ7.5.4D 和 ab11575) 及 丙 二 醛(malondialdehyde,MDA)、羰基化蛋白、超氧化物歧化酶(superoxidase dismutase,SOD)和过氧化氢酶(catalase,CAT) 检测试剂盒均购自Abcam(上海)公司,二抗(Alexa Fluor 546 标记的驴抗鼠和Alexa Fluor 488 标记的驴抗山羊抗体)和DAPI 购自赛默飞(上海)公司,K-H 液购自麦凯吉(北京) 公司。血糖仪和试纸购自罗氏诊断(上海)公司,体外肌肉收缩力检测系统购自奥尔科特(上海)公司。
1.2 大鼠GDM 模型构建大鼠GDM 模型构建方法参照文献[7]。实验开始前3 d,每天记录大鼠的体质量并测量血糖。实验开始前将体质量差异较大和血糖水平异常(7 mmol·L-1以上)大鼠剔除。剩余大鼠按雌雄2∶1 比例进行合笼交配,次日清晨观察到阴栓即认定交配成功,并标记为妊娠第0 天。将妊娠成功的44 只母鼠随机分为GDM 组和正常对照组,每组22 只。GDM 组孕鼠于妊娠成功后的12 h 后,单次腹腔注射1% STZ 溶液(35 mg·kg-1),正常对照组孕鼠单次腹腔注射相同剂量的0.1 mmol·L-1柠檬酸-柠檬酸钠缓冲液。大鼠妊娠第4 天,经眼眶取血,测量空腹血糖水平在11.1 mmol·L-1以 上,随机血糖 在17.9 mmol·L-1以上,尿糖>,表明GDM 造模成功。共有18 只大鼠成功构建GDM 模型。
1.3 子代大鼠膈肌收缩力测量体外膈肌收缩力的测量参考既往研究[8]。分娩后,从每只母鼠的子代中随机挑选1 只新生子代大鼠(正常对照组22 只,GDM 组18 只),置于天平上称质量后处死,取出完整膈肌组织并测量湿质量。 剪取左侧肋弓下膈肌组织,修剪成5 mm×3 mm 的肌条。固定夹固定肌条两端,垂直放置于27 ℃、95%O2和5%CO2混合气体持续氧合的K-H 液中平衡30 min。在刺激电压为10 V、波长2 ms 和刺激间隔2 min 条件下不断调整肌条的长度,从而获取肌条的最佳初长度。膈肌的最佳初长度为膈肌在以上刺激条件下达到最大紧张收缩力时的长度。最大紧张收缩力即为最佳初长度下的膈肌收缩力,测量3 次取平均值。将刺激电压设为15 V,其他条件不变,在最佳初长度下获取最大强直收缩力,取3 次测量平均值。在其他条件不变的情况下,改变刺激频率,依次获取膈肌在20、40、60、80、100 和120 Hz 时的收缩力。在刺激电压10 V、刺激间隔2 min、波长2 ms、刺激频率60 Hz、串数120、串间隔670 ms、刺激时间330 ms 和串长16.5 的条件下行疲劳检测,获取第1个和第120个波形的收缩力,2 个波形收缩力减少的百分比即为疲劳指数。实验结束时,测定肌条在最佳初长度下的物理长度(cm)和肌条的湿质量(g),按照肌肉密度为1.06 g·cm-3,每个肌条在最佳初长度下的横截面积(cm2)=湿质量/密度/肌条长度。
1.4 子代大鼠膈肌纤维横截面积测量将取材的子代大鼠左侧肋弓下膈肌组织用OCT 包埋,冰冻切片机切成8 μm 厚的切片,冰甲醇固定30 s,PBS洗 涤。 MY-32 一 抗、 NOQ7.5.4D 一 抗 和 抗Laminin 抗体按1∶300 比例双色免疫荧光标记,湿盒中4℃避光过夜。PBS 洗涤,二抗室温下孵育1 h后,洗涤,加入DAPI,封片。荧光显微镜下拍照,随机选取5 个不同视野,采用ImageJ 软件计算Ⅰ型和Ⅱ型膈肌纤维的横截面积,每只大鼠至少统计200 条Ⅰ型和Ⅱ型膈肌纤维。
1.5 子代大鼠膈肌组织形态表现和超微结构观察取子代大鼠左侧肋弓下膈肌组织,部分膈肌沿肌纤维横切面修剪后用4%多聚甲醛固定,常规脱水、透明、浸蜡包埋后行HE 染色,在普通光学显微镜下观察膈肌组织形态表现。部分膈肌用2.5%戊二醛固定处理后,行超薄切片,3%醋酸铀-枸橼酸铅双染色,透射电镜下观察膈肌超微结构。
1.6 子代大鼠膈肌组织中氧化应激指标检测称取取材的子代大鼠左侧肋弓下膈肌组织50 mg,加入800 μL RIPA 裂解液,冰上匀浆后,采用低温离心机1 500 r·min-1离心5 min 后提取上清液。根据检测试剂盒的说明,采用比色法检测上清液中的MDA 和羰基化蛋白水平及SOD 和CAT 活性。
1.7 统计学分析采用SPSS 19.0 统计软件进行统计学分析。采用Shapiro-Wilk 检验验证各组数据的正态分布情况。子代大鼠体质量,膈肌湿质量,膈肌收缩力,膈肌疲劳指数,膈肌纤维横截面积,膈肌组织中MDA 和羰基化蛋白水平及SOD 和CAT 活性均符合正态分布,以表示,2 组间样本均数比较采用两独立样本t检验。以P<0.05为差异有统计学意义。
2 结 果
2.1 2 组子代大鼠体质量、膈肌湿质量和收缩力GDM 组子代大鼠体质量[(8.44±0.11) g] 和膈肌湿质量[(0.45±0.01) g] 与正常对照组[(8.39±0.10)和(0.44±0.01)g]比较差异均无统计学意义(P>0.05)。与正常对照组比较,GDM 组子代大鼠膈肌最大紧张收缩力、最大强直收缩力和不同刺激频率下的膈肌收缩力均明显降低(P<0.05),膈肌疲劳指数明显升高(P<0.05)。见表1。
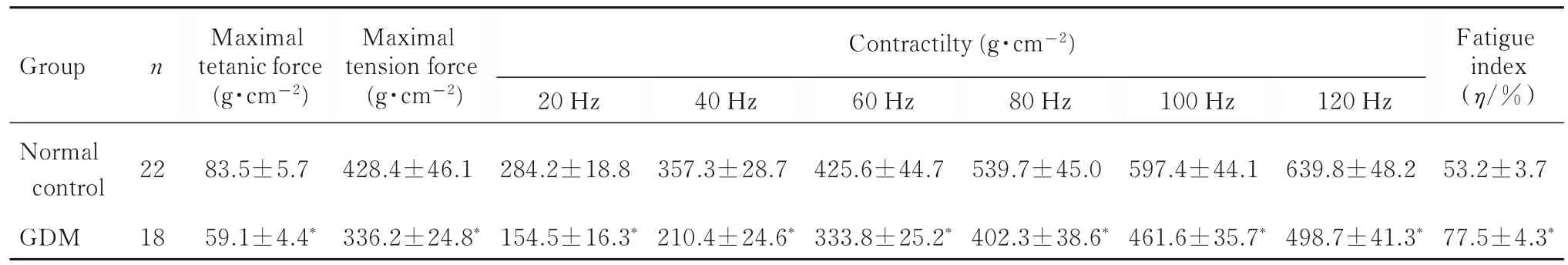
表1 2 组子代大鼠膈肌收缩力和膈肌疲劳指数Tab.1 Diaphragmatic contractile forces and fatigue indexes of offspring rats in two groups (x±s)
2.2 2组子代大鼠膈肌纤维横截面积GDM 组子代大鼠Ⅰ型和Ⅱ型膈肌纤维横截面积[(613±85) 和 (1 406±159) μm2] 较 正 常 对 照 组[(859±107)和(1 748±183)μm2]明显缩小,差异均有统计学意义(P<0.05)。见图1。
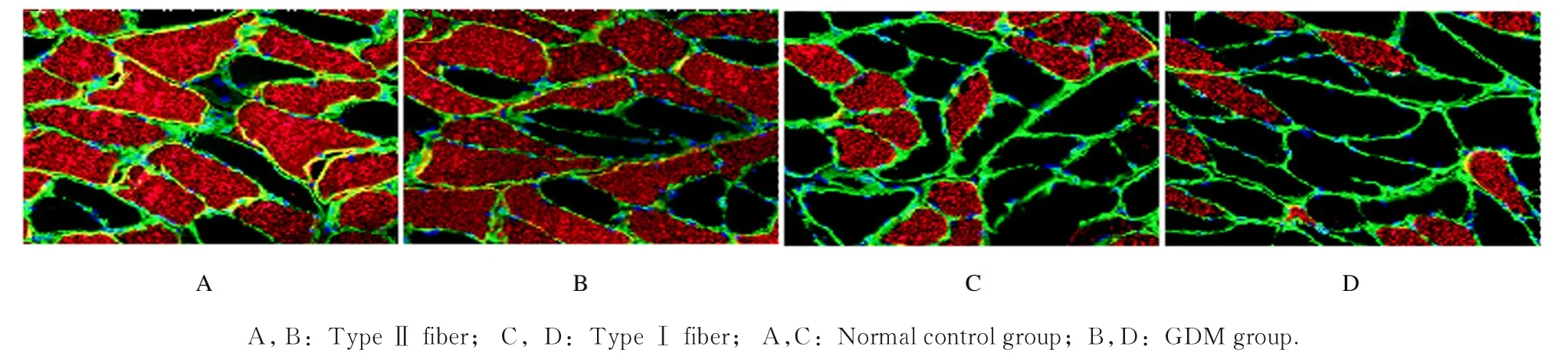
图1 2 组子代大鼠膈肌纤维免疫荧光检测结果(×400)Fig.1 Results of immunofluorescence detection of diaphragm fibers of offspring rats in two groups(×400)
2.3 2 组子代大鼠膈肌组织形态表现和超微结构HE 染色结果显示:2 组子代大鼠膈肌纤维结构完整,排列整齐、致密,膈肌细胞无明显肿胀,细胞核染色清晰。见图2。
透射电镜下观察,正常对照组子代大鼠膈肌纤维排列整齐,明暗带及“Z”线清晰;线粒体结构完整,嵴成隔板状,清晰可见,排列规则。GDM 组子代大鼠膈肌纤维排列紊乱,明暗带及“Z”线模糊或断裂;线粒体外膜模糊,形态不规则,嵴消失呈空泡化,出现凋亡表现。见图3。
2.4 2 组子代大鼠膈肌组织中MDA 和羟基化蛋白水平及SOD 和CAT 活性与正常对照组比较,GDM 组子代大鼠膈肌组织中MDA 和羰基化蛋白水平均明显升高(P<0.05),SOD 和CAT 活性均明显降低(P<0.05)。见表2。
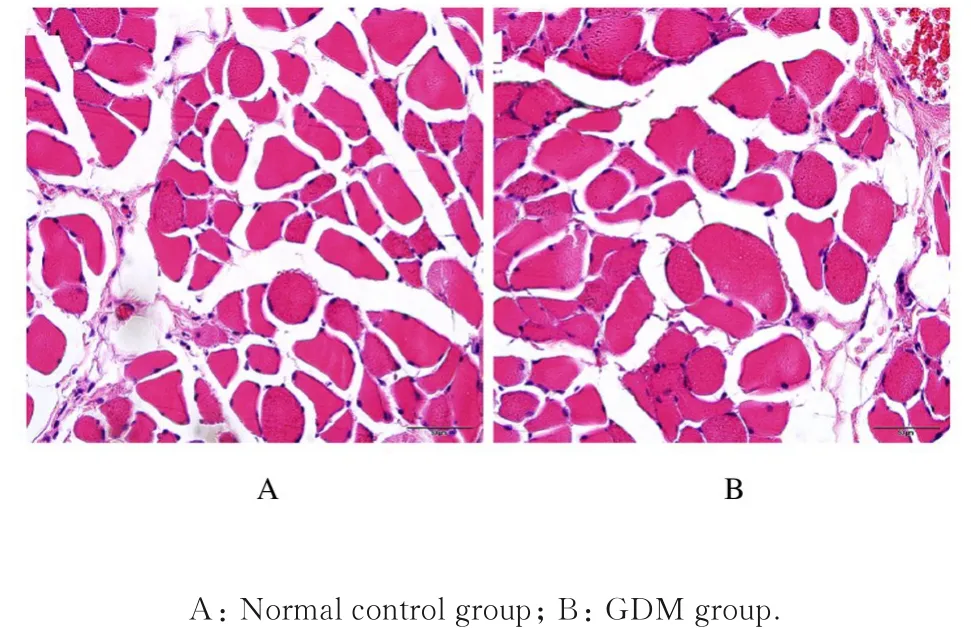
图2 2 组子代大鼠膈肌组织形态表现(HE,×400)Fig.2 Morphology of diaphragm tissue of offspring rats in two groups(HE, ×400)
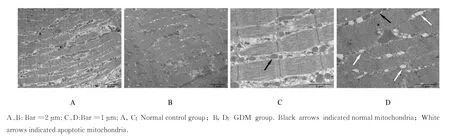
图3 2 组子代大鼠膈肌组织超微结构Fig.3 Ultrastructures of diaphragm tissue of offspring rats in two groups

表2 2 组子代大鼠膈肌组织中MDA 和羟基化蛋白水平及SOD 和CAT 活性Tab.2 Levels of MDA and carbonylated protein and activities of SOD and CAT in diaphragm tissue of offspring rats in two groups (x±s)
3 讨 论
本研究结果表明:GDM 母鼠高血糖状态会影响新生子代大鼠的膈肌收缩功能,并引起明显的形态学改变。GDM 是产科的常见疾病之一,在我国,GDM 的发病率为1%~5%。近年来,随着生活水平的提高和生活习惯的改变,我国GDM 发病率一直呈上升趋势[9]。对于孕妇,妊娠期间持续的高血糖状态可能导致母体出现妊高症、胎膜早破、羊水过多及早产等并发症。此外,母体内的高血糖可以通过胎盘进入胎儿体内,导致胎儿出现高血糖和继发性的高胰岛素血症。胰岛素作为胎儿的主要生长激素,过多的胰岛素会导致胎儿过度生长[10]。已有证据[4]表明:高血糖和高胰岛素血症引起胎儿心脏超微结构和功能的改变。膈肌是人体的主要呼吸肌,贡献人体70%~85%的呼吸做功。新生儿的呼吸频率为40~60 min-1,远高于成年人的呼吸频率(16~24 min-1),因此新生儿在完成通气性气体交换的过程中对膈肌的依赖程度远远高于成年人。一旦胎儿期间膈肌功能发育异常,会导致新生儿期出现呼吸功能不全,可能进一步引起缺氧、呼吸衰竭甚至死亡[11]。本研究首次揭示了GDM 对新生子代大鼠的膈肌功能的影响,结果显示:妊娠期高血糖状态会导致新生子代大鼠的膈肌收缩力降低和膈肌抗疲劳能力下降。肌肉的收缩力与肌纤维的直径有关[12]。本研究结果显示:伴随膈肌收缩力的降低,妊娠期高血糖状态也会导致新生子代大鼠膈肌的Ⅰ型和Ⅱ型膈肌纤维横截面积缩小,说明高血糖状态会影响胎儿期间膈肌纤维的发育,导致新生子代膈肌纤维的直径相对较小,进而影响膈肌的收缩力。肌肉的收缩力与肌小结的结构有关[13]。本研究进一步观察子代大鼠膈肌组织形态表现和超微结构,结果显示:妊娠期高血糖不影响膈肌纤维的大体形态,但可造成膈肌纤维排列紊乱,明暗带及“Z”线模糊或断裂,线粒体嵴消失呈空泡化,出现凋亡改变,说明高血糖可直接影响膈肌纤维肌小结的改变,从而造成膈肌收缩力降低。
目前,临床研究和动物实验研究结果[14-15]均表明:氧化应激水平升高是引起膈肌功能异常的关键因素。本研究通过检测子代大鼠膈肌组织中MDA 和羰基化蛋白水平的变化来反映氧化应激水平的高低,结果显示:GDM 组子代大鼠膈肌组织中MDA 和羰基化蛋白水平明显高于正常对照组,说明妊娠期的高血糖状态会导致新生子代大鼠膈肌组织高氧化应激水平。而氧化应激引起膈肌收缩力降低的机制,目前普遍认为膈肌中的氧化应激水平的升高通过激活膈肌中一系列蛋白水解酶体系,如蛋白酶(calpain)系统、含半胱氨酸的天冬氨酸蛋白水解酶3(caspase-3)系统、溶酶体系统以及泛素-蛋白酶体系统,共同引起蛋白水解是其主要机制[16-18]。在蛋白水解加强的同时,膈肌的蛋白合成受到部分抑制,导致肌纤维出现萎缩,进而导致膈肌收缩力降低。而正常体内活性氧类物质的生成和清除处于动态平衡。氧化应激是指体内氧化与抗氧化作用失衡的一种状态,氧化权重加大,导致中性粒细胞炎性浸润,蛋白酶分泌增加,产生大量氧化中间产物。在氧化权重加大的同时,抗氧化酶体系,如SOD 和CAT,可以分解活性氧类物质以减少氧化应激损伤[19-20]。本研究结果表明:GDM 组子代大鼠膈肌组织中SOD 和CAT 活性明显低于正常对照组,说明妊娠期高血糖状态会抑制抗氧化酶的活性或导致抗氧化酶体系的过度消耗。本研究结果反映了妊娠期母体高血糖状态会导致胎儿或新生子代膈肌持续的氧化氧化应激损伤,而氧化应激损伤可能是导致新生子代膈肌功能异常的原因之一。
综上所述,GDM 可引起新生子代大鼠膈肌收缩力降低、纤维横截面积减小和膈肌超微结构的改变,而氧化应激水平升高可能是膈肌功能发生异常的原因之一。
猜你喜欢
中国临床医学影像杂志(2022年6期)2022-07-26
中国临床医学影像杂志(2022年5期)2022-07-26
国际放射医学核医学杂志(2021年10期)2021-02-28
现代检验医学杂志(2016年3期)2016-11-15
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
广西林业科学(2016年3期)2016-03-16
中国病理生理杂志(2015年8期)2015-12-21
中国医疗美容(2015年4期)2015-04-27
