
五倍子酸通过调节人肝癌HepG-2 细胞线粒体氧化磷酸化对细胞凋亡的诱导作用
2021-01-31 08:03张海英张文馨赵文静
吉林大学学报(医学版) 2021年1期
张海英, 张文馨, 赵文静, 李 伟
(1. 吉林大学基础医学院 病理生物学教育部重点实验室,吉林 长春 130021;2. 吉林省肝胆病医院中心实验室,吉林 长春 130062)
哺乳动物细胞可通过线粒体氧化磷酸化(oxidative phosphorylation,OXPHOS) 和糖酵解2 种糖代谢方式供应能量ATP,常氧情况下,正常细胞糖代谢主要由线粒体OXPHOS 产生ATP 以提供代谢活动所需要的能量。WARBURG 等[1]认为:肿瘤细胞由于线粒体呼吸缺陷,即使在有氧环境中也通过糖酵解供应大部分ATP。但近期研究[2-4]表明:多数肿瘤细胞线粒体是完整的,即使有些细胞具有更高的糖酵解,但仍保持线粒体呼吸,在某些癌症(包括白血病、淋巴瘤、胰腺导管腺癌、高OXPHOS 亚型黑色素瘤、子宫内膜癌和高侵袭性肿瘤等)中线粒体OXPHOS 发挥重要作用,OXPHOS 是抗肿瘤治疗的新靶点。五倍子酸(3,4,5-三羟基苯甲酸,gallic acid,GA) 也称为没食子酸或棓酸,是一种天然多酚类化合物,已证实其对多种肿瘤细胞系具有抗肿瘤活性[5-8]。但GA 对肝癌细胞线粒体OXPHOS 的抑制作用尚未见报道。本研究以人肝癌HepG-2 细胞为模型,首次采用Seahorse Extracellular Analyzer 细胞能量代谢分析仪检测GA 作用下HepG-2 细胞量代谢动态全过程,分析细胞线粒体膜电位(mitochondrial membrane potential,MMP)、线粒体细胞色素C(cytochrome C,CytC) 和细胞中ATP 水平的变化,从细胞能量代谢和与之密切相关的细胞器线粒体出发,探讨GA 抗肝癌的作用机制。
1 材料与方法
1.1 细胞、主要试剂和仪器人肝癌HepG-2 细胞(江苏凯基生物技术股份有限公司),GA (美国Chromadex 公司),线粒体膜电位检测试剂盒、Annexin V-FITC/PI 细胞凋亡检测试剂盒、线粒体分离试剂盒和胞浆蛋白抽提试剂盒(南京碧云天生物技术有限公司),CytC 单克隆抗体和β-actin 抗体(美国SAB 公司),化学发光检测试剂盒(上海天能科技有限公司),胎牛血清和DMEM 培养基(美国Gibco 公司),线粒体压力检测试剂盒(美国Seahorse Bioscience 公司),ATP 检测试剂盒(南京建成生物工程研究所)。二氧化碳培养箱(美国Thermo Fisher Scientific 公司),全波长多功能酶标仪(美国Perkinelmer 公司),高速冷冻离心机(上海安亭科学仪器厂),倒置荧光显微镜(日本Olympus 公司),电泳仪和半干转印槽转膜仪(美国Bio-Rad 公司),流式细胞仪(美国BD 公司)。
1.2 MTT 法检测HepG-2 细胞存活率参照文献[9]方法,HepG-2 细胞以 每 孔100 μ L (1×105mL-1) 接种到96 孔细胞培养板,培养24 h 后分为对照组和不同浓度GA 组(加入终浓度0、3.125、 6.250、 12.500、 25.000、 50.000 和100.000 mg·L-1GA[10]),每个浓度设置4 个复孔,作用24、48 和72 h 后,每孔加入20 μL MTT溶液,孵育4 h 后终止培养弃去培养液,每孔加入150 μL DMSO 充分溶解紫色结晶物,酶标仪490 nm 下测定吸光度(A)值,计算细胞存活率。细胞存活率=(实验组A 值-空白对照组A 值)/(对照组A 值-空白对照组A 值)×100%。
1.3 能量代谢分析仪检测细胞线粒OXPHOS 水平对数生长期HepG-2 细胞接种于Seahorse XFp专用板(8 000 个/孔),培养24 h 后,分为对照组及6.25、 12.50 和25.00 mg·L-1GA 组,分别加入 含0、 6.25、 12.50 和25.00 mg·L-1GA 的DMEM 培养基作用24 h,更换能量分析专用培养液,按照线粒体压力测定试剂盒说明书配制试剂,能量代谢分析仪实时检测细胞线粒体耗氧率(oxygen consumption rate,OCR),代表细胞线粒体OXPHOS 活性,并根据OCR 值分析各组HepG-2 细胞的基础有氧呼吸、最大有氧呼吸能力、有氧呼吸储备能力和ATP 产生量。
1.4 流式细胞术检测细胞MMP每孔1×106个HepG-2 细胞接种于6 孔细胞培养板,培养24 h 后分组同“1.3”,分别加入含(0、6.25、12.50 和25.0 mg·L-1)GA 的DMEM 培养基作用12 h,收集各组细胞按照Rhodamine123 染色试剂盒操作,激发波长505 nm,发射波长530 nm,流式细胞仪测定线粒体内Rhodamine123 的荧光强度,以强荧光细胞百分率表示MMP。
1.5 比色法检测细胞中ATP 水平细胞分组及处理同“1.4”,离心收集各组细胞,加入提取液,冰浴超声波破碎5 min,4 ℃、12 000 r·min-1离心3 min,取上清,置冰上检测。严格按照ATP 水平测定试剂盒操作,分光光度计在波长636 nm 处测定各管A 值。ATP 水平(μmol·g-1prot)=(测定A 值-对照A 值) / ( 标 准 A 值 - 空 白A 值)×标准品浓度(1×103μmol·L-1)× 样本测定前稀释倍数/待测样本蛋白浓度(g·L-1)。
1.6 Western blotting 法检测细胞胞浆和线粒体中CytC 蛋白表达量细胞分组同“1.4”,收集GA处理48 h 的HepG-2 细胞,按照线粒体分离试剂盒操作制备线粒体裂解液,按照细胞胞浆蛋白抽提试剂盒操作制备胞质部分裂解液,BCA 法测定蛋白浓度。取20 μg 蛋白样品上样,经SDS-PAGE 电泳分离后,半干转蛋白至硝酸纤维素膜(NC) 上,5%脱脂奶粉室温封闭3 h,一抗4℃孵育过夜,辣根过氧化物酶标记的二抗室温孵育2 h,ECL 试剂进行化学发光。胶片于暗室曝光后,扫描并观察结果。
1.7 吖啶橙染色观察细胞形态表现细胞分组同“1.4”,收集GA 处理48 h 的HepG-2 细胞,制成细胞悬液,调整细胞浓度为1×107mL-1,95 μL 细胞悬液加入5 μL 吖啶橙储存液(0.1 g·L-1),染色15 min 后取1 滴细胞悬液点于洁净玻片上,盖玻片封片,荧光显微镜下观察细胞形态表现。
1.8 流式细胞术检测细胞凋亡率每孔1×106个HepG-2 细胞接种于6 孔细胞培养板,24 h 后分为对照组及6.25、12.50 和25.00 mg·L-1组,更换含GA 0、 6.25、 12.50 和 25.00 mg·L-1GA 的DMEM 培养基作用48 h,离心收集细胞,按照Annexin Ⅴ-FITC/PI 细胞凋亡检测试剂盒说明书操作。每个样品加入5 μL Annexin V 溶液和5 μL PI 溶液避光孵育15 min,最后各加入400 μL PBS缓冲液,流式细胞仪检测细胞凋亡。早期细胞凋亡率=早期凋亡细胞数/总细胞数×100%,晚期细胞凋亡率=晚期凋亡细胞数/总细胞数×100%。
1.9 统计学分析采用SPSS17.0 统计软件进行统计学分析。各组HepG-2 细胞存活率、基础有氧呼吸、最大有氧呼吸能力、有氧呼吸储备能力、ATP 产生量、MMP 和ATP 水平以及细胞凋亡率均符合正态分布,以表示,多组间样本均数比较采用单因素方差分析,组间样本均数两两比较采用SNK-q法。以P<0.05 为差异有统计学意义。
2 结 果
2.1 各组HepG-2 细胞存活率对照组HepG-2 细胞生长活跃,经不同浓度GA 处理24、48 和72 h后,各剂量GA 组HepG-2 细胞存活率均不同程度降低(P<0.05 或P<0.01),并呈明显的时间和浓度依赖性。见图1。
2.2 各组HepG-2 细胞线粒体OXPHOS与对照组比较,不同浓度GA 组HepG-2 细胞基础有氧呼吸、最大有氧呼吸能力、有氧呼吸储备能力和ATP 产生量均明显降低(P<0.05 或P<0.01)。见图2。
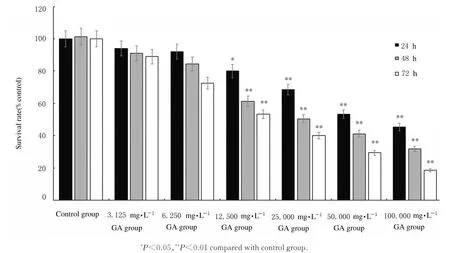
图1 GA 作用不同时间各组HepG-2 细胞存活率Fig.1 Survival rates of HepG-2 cells in various groups after treated with GA for different time
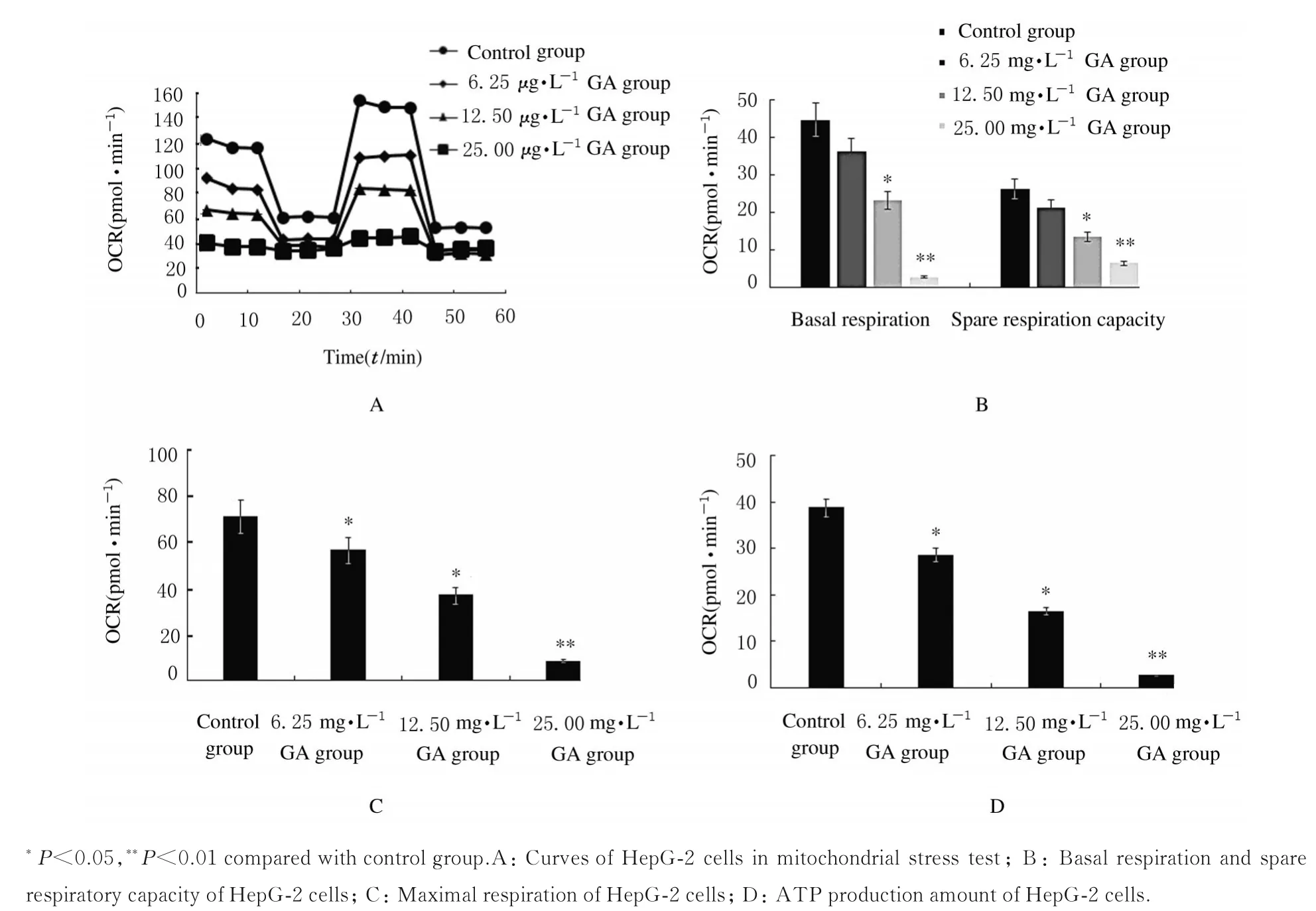
图2 各组HepG-2 细胞线粒体OXPHOSFig.2 Mitochondrial OXPHOS of HepG-2 cells in various groups
2.3 各组HepG-2 细胞MMP 和ATP 水平GA 作用24 h,HepG-2 细胞中强荧光细胞百分率逐渐减少。6.25、12.50 和25.00 mg·L-1GA 组强荧光细胞 百 分 率 即MMP 分 别 为(73.12±4.61)%、(65.49±4.32)%和(38.64±5.93)%,与对照组[(94.38±2.18)%] 比较均明显降低(P<0.05),见图3。与对照组比较,不同浓度GA 组细胞 中ATP 水 平 均 明 显 降 低(P<0.05 或P<0.01),6.25、12.50 和25.00 mg·L-1GA 组 细胞中ATP 水平分别降低至61.11%、45.56% 和30.24%,见图4。
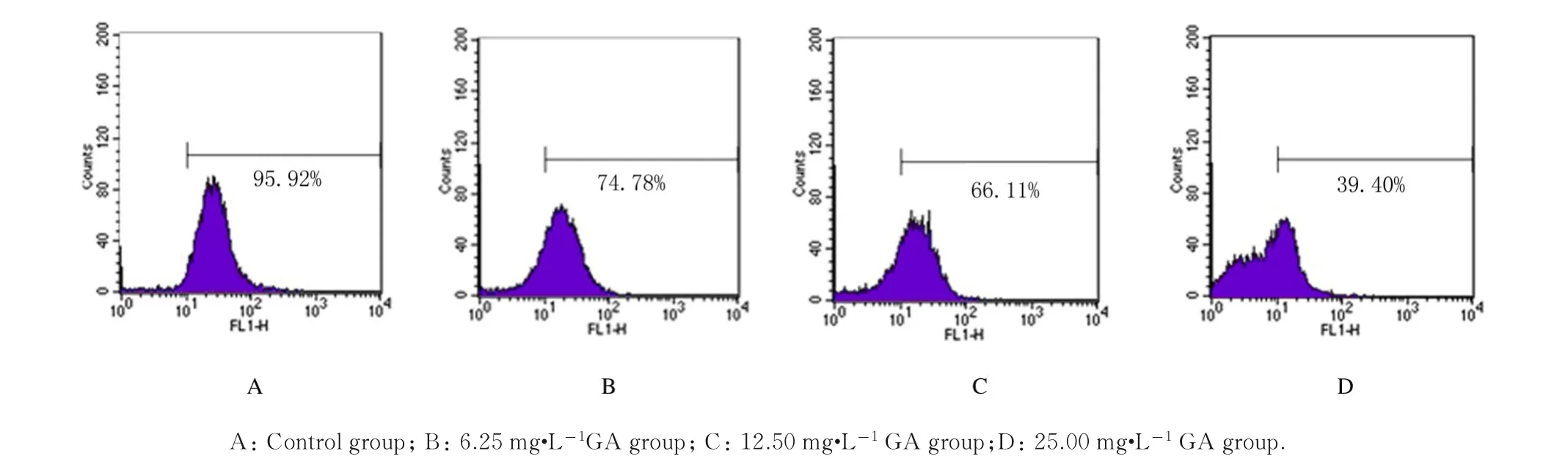
图3 各组HepG-2 细胞MMPFig.3 MMP of HepG-2 cells in various groups
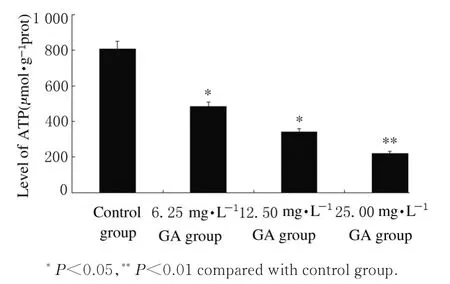
图4 各组HepG-2 细胞ATP 水平Fig. 4 Levels of ATP in HepG-2 cells in various groups
2.4 各组HepG-2 细胞胞浆和线粒体中Cyt C 表达量对照组HepG-2 细胞胞浆无Cyt C 特异条带,不同浓度GA 组细胞胞浆均检测到Cyt C 表达,随着GA 浓度的增加胞浆中Cyt C 表达量明显增加;各组细胞线粒体均有Cyt C 特异条带出现,不同浓度GA 组线粒体中Cyt C 表达量随着GA 剂量的增加呈降低趋势。见图5。
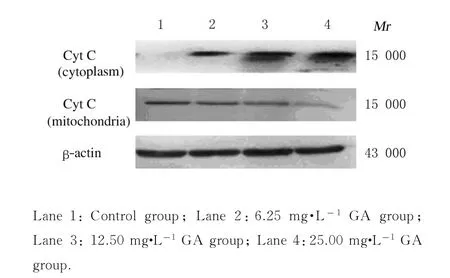
图5 各组HepG-2 细胞胞浆和线粒体中Cyt C 蛋白表达电泳图Fig. 5 Electrophoregram of expressions of Cyt C protein in cytoplasm and mitochondria of HepG-2 cells in various groups
2.5 各组HepG-2 细胞凋亡形态表现及凋亡率对照组HepG-2 细胞形态饱满,细胞质呈均匀弥散绿色荧光,细胞核形态规则; GA 处理48 h 后不同浓度GA 组HepG-2 细胞体积缩小,细胞核碎裂,染色质凝集,呈凋亡细胞的形态学变化(图6)。进一步采用Annexin V-FITC/PI 双染定量检测细胞 凋 亡 率,GA 作 用48 h 后,6.25、12.50 和25.00 mg·L-1GA 组 早 期 细 胞 凋 亡 率 分 别 为(10.8±0.8)% 、(18.7±0.7)% 和 (26.9±0.8)%,与对照组[(0.2±0.1)%]比较明显升高(P<0.05);晚期细胞凋亡率分别为(1.8±0.4)%、(11.2±0.6)%和(20.1±0.5)%,与对照组[(0.3±0.1)%]比较明显升高(P<0.05)。见图7。

图6 吖啶橙染色观察各组HepG-2 细胞形态表现(×100)Fig.6 Morphology of HepG-2 cells in various groups observed by acridine orange staining(×100)
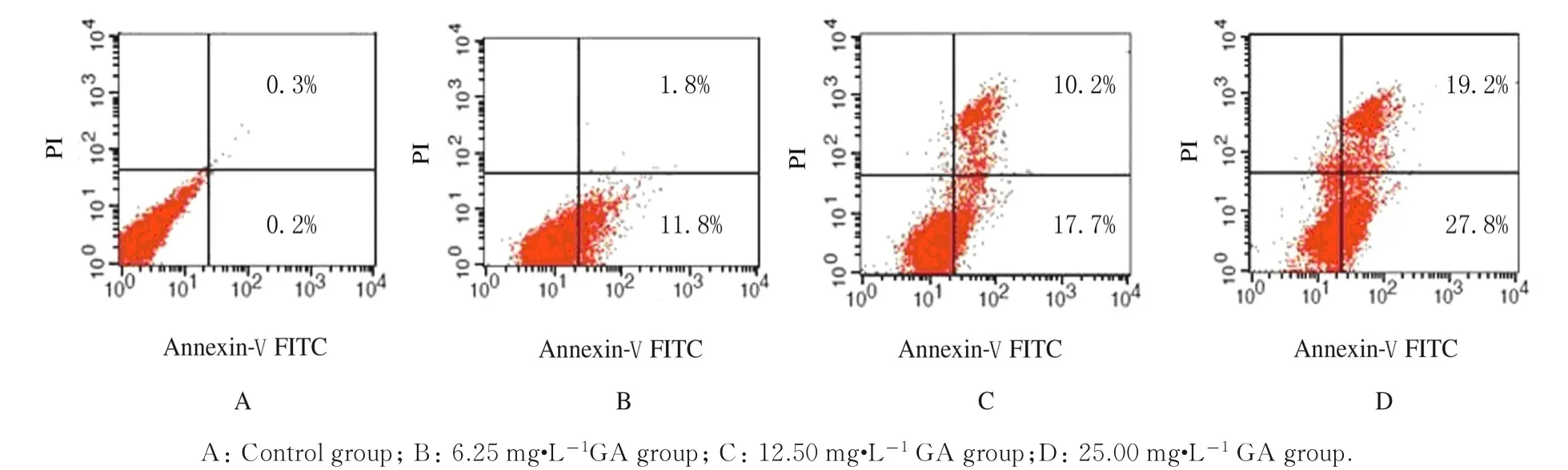
图7 流式细胞术检测各组HepG-2 细胞凋亡率Fig.7 Apoptotic rates of HepG-2 cells in various groups detected by flow cytometry
3 讨 论
全球50% 以上的新发和死亡肝癌患者在中国[11]。肝癌起病隐匿,外科手术是首选治疗方法,但能获得手术切除机会的患者仅占20%~30%[12]。肝癌患者术后5 年复发率高达50%~70%,存活率仅为20%~30%[13]。因此研发高效、低毒的抗肝癌药物十分迫切[14-15]。本研究利用人肝癌HepG-2细胞,从细胞增殖、线粒体OXPHOS 抑制和细胞凋亡等方面观察GA 对HepG-2 的抗肿瘤作用及潜在机制。
近年来,多个科研团队[16-18]将OXPHOS 抑制剂用于肿瘤治疗均取得了满意的疗效。ZHANG 等[16]研究发现:OXPHOS 抑制剂IACS-010759 在体内体外可明显抑制伊布替尼耐药的套细胞淋巴瘤细胞的生长。ZHAO 等[17]研究发现:维生素E 衍生物ESeroS GS 可通过下调己糖激酶2(hexokinase 2,HK2)及具有电子传递功能的复合物Ⅱ/Ⅲ/Ⅳ(complexⅡ/Ⅲ/Ⅳ) 等的表达,降低乳腺癌细胞MMP,同时抑制乳腺癌细胞糖酵解和OXPHOS,从而抑制肿瘤细胞的增殖、迁移和侵袭;范霞霞等[18]研究表明:天然二萜衍生物Jar-TTA 通过双重抑制糖酵解和线粒体OXPHOS诱导食管癌细胞凋亡,其机制可能与Jar-TTA 降低食管癌细胞MMP 进而干扰线粒体OXPHOS 功能,以及下调葡萄糖转运蛋白4(glucose transporter 4,GLIT4) 和乳酸脱氢酶A(L-actate dehydrogenase A chain,LDHA)而抑制葡萄糖摄取和糖酵解进程有关。线粒体内膜(inner mitochondrial membrane,IMM) 是OXPHOS 产生ATP 的部位,线粒体基质中的三羧酸循环利用葡萄糖的代谢产物产生烟酰胺 腺 嘌 呤 二 核 苷 酸 (nicotinamide adenine dinucleotide,NADH),并将其提供给位于线粒体内膜的电子传递链(electron transport chain,ETC),线粒体ATP 合成酶利用ETC 产生的质子梯度作为驱动力催化ADP 磷酸化生成ATP[19-20]。本研究中首先采用MTT 法检测GA 对HepG-2 细胞增殖的影响,结果显示:在体外GA 可有效抑制HepG-2 细胞增殖;细胞能量代谢分析仪检测GA可明显抑制HepG-2 细胞线粒体OXPHOS 活性,与对照组比较,GA 处理组HepG-2 细胞基础有氧呼吸、最大有氧呼吸能力、有氧呼吸储备能力和ATP 产生量均明显降低。
IMM 的完整和非渗透性保证了线粒体功能的正常[21],MMP 反应了沿着电子传递链电子传递而形成的质子梯度,是线粒体功能完整性的指标,其完整性主要由ETC 和ATP 合酶维持[22]。本研究结果显示:GA 能明显降低HepG-2 细胞MMP,减少ATP 产生。HepG-2 细胞MMP 下降,导致线粒体通透性增加,存在于线粒体中的Cyt C 释放入胞浆,Cyt C 启动细胞凋亡线粒体信号转导通路,最终激活半胱氨酸蛋白酶(cysteinyl aspartate specific proteinase,caspase)级联反应诱导HepG-2细胞凋亡[23-25]。本研究中吖啶橙染色和流式细胞仪检测结果显示: 不同浓度GA 在体外可诱导HepG-2 细胞凋亡,不同浓度GA 作用HepG-2 细胞48 h 后,早期细胞凋亡率达到和晚期凋亡细胞率达到均明显提高。
综上所述,本研究将细胞能量代谢与抑制肿瘤细胞增殖、诱导肿瘤细胞凋亡相关联,阐明GA 可能通过降低HepG-2 细胞MMP,抑制线粒体OXPHOS 功能,诱导肿瘤细胞凋亡。GA 是否对线粒体中具有电子传递功能的complex Ⅱ/Ⅲ/Ⅳ具有调控作用需进一步研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
中老年保健(2022年2期)2022-08-24
中老年保健(2022年4期)2022-08-22
建材发展导向(2021年11期)2021-07-28
当代水产(2020年10期)2020-03-17
当代水产(2019年8期)2019-10-12
中华老年多器官疾病杂志(2016年8期)2016-05-14
中国男科学杂志(2016年9期)2016-03-20
中国体育教练员(2015年2期)2015-04-16
现代检验医学杂志(2015年2期)2015-02-06
