
外源诱导姬松茸麦角甾醇的合成
2021-01-19 02:37:08范秀芝姚芬殷朝敏史德芳高虹
现代食品科技 2021年1期
范秀芝,姚芬,殷朝敏,史德芳,高虹
(湖北省农业科学院农产品加工与核农技术研究所,湖北武汉 430064)
姬松茸(Agaricus blazei)又名巴氏蘑菇、巴西蘑菇[1],属于担子菌门、伞菌目、蘑菇科、蘑菇属(Agaricus)。姬松茸味道鲜美,含有丰富的蛋白质、人体必需氨基酸、矿物质和微量元素等营养成分。近年来,随着国内外对姬松茸研究的不断深入,发现它不仅是一种美味的食用菌,还在抗肿瘤、抗氧化、抗衰老、抑菌、降血糖、抗血栓等方面效果奇特[2],尤其是其抗肿瘤作用居食(药)用菌首位[3]。
2001年,Takaku等从干的姬松茸子实体中分离到一种脂类物质,通过研究发现该物质能直接抑制肿瘤生长,经核磁共振和质谱分析后确定此物质为麦角甾醇[4]。2011年,高虹等[5]发现姬松茸菌丝体醇提物中的主要抗肿瘤活性成分也是麦角甾醇,并经试验证实麦角甾醇是通过抑制肿瘤新生血管的形成来抑制肿瘤生长。已知抑制新生血管形成是抑制肿瘤的潜在治疗手段[6],因此,这一发现为姬松茸在医药领域的应用提供了依据。但目前姬松茸麦角甾醇含量相对较低,子实体中含量仅为1.00~2.00 mg/g[7],且姬松茸子实体的栽培周期较长,栽培过程需要消耗大量人力和物力,产量也不高,这些问题限制了姬松茸子实体作为原料用于其麦角甾醇产品的开发及应用。
外源诱导物,又称诱导子(elicitor molecules)、激发子(elicitor),是当生物受到胁迫时产生的多种能够响应胁迫并诱导防御基因表达,以增强抗逆性的一类特殊化合物的总称[8],如茉莉酸类(jasmonates,JAs)和水杨酸(salicylic acid,SA)等。大量研究表明,JAs和SA可诱导酶和防御蛋白的活性水平,导致次级代谢物质的产生和积累,被认为是非常有效的诱导子[9,10]。近年来,JAs的茉莉酸甲酯(methyl jasmonate,MeJA)和水杨酸(salicylic acid,SA)在大型真菌代谢产物诱导应用中初见成效。已有的研究主要集中在灵芝(Ganoderma lucidum)三萜[11]、桑黄(Phellinus igniarius)黄酮[12]和猴头菌(Hericium erinaceus)麦角甾醇[13]产量提高及诱导机制的研究中。
因此,本研究借助液体培养的优势,通过外源诱导物的添加提高菌丝中麦角甾醇产量,并通过诱导条件下参与麦角甾醇合成基因的表达分析,探析姬松茸麦角甾醇合成的分子机理及其调控机制,为利用基因工程手段构建高产麦角甾醇工程菌株提高麦角甾醇的生物合成量,以及开发保健食品及功能配方食品奠定基础,同时为进一步全面阐述姬松茸中次生活性物质代谢合成调控机制提供理论指导。
1 材料和方法
1.1 材料与试剂
姬松茸菌株 JSR3购自华中农业大学菌种实验中心,保藏于湖北省农业科学院农产品加工与核农技术研究所。
麦角甾醇标准品购自上海源叶生物科技有限公司;茉莉酸甲酯、水杨酸、色谱级甲醇等购自 Sigma公司;氢氧化钾、无水乙醇、正己烷等试剂均为分析纯。
1.2 培养基及培养条件
固体完全培养基(CYM):葡萄糖20.00 g,蛋白胨2.00 g,酵母膏2.00 g,KH2PO40.46 g,K2HPO41.00 g,MgSO4·7H2O 0.50 g,琼脂20.00 g,蒸馏水 1.00 L。
JSR3菌株接种到CYM固体培养基,于25 ℃,避光培养5~7 d进行活化。
1.3 麦角甾醇诱导合成条件优化
1.3.1 茉莉酸甲酯(methyl jasmonate,MeJA)和水杨酸(salicylic acid,SA)添加浓度筛选
(1)MeJA添加浓度筛选
分别向100 mL(250 mL三角瓶)液体CYM培养基中加入不同体积的无菌MeJA溶液使其终浓度分别为 60.00、70.00、80.00、90.00、100.00 μmol/L。活化的姬松茸菌丝用 8 mm打孔器打孔,等量接种到CYM液体培养基中,25 ℃静止培养2 d后于120 r/min条件下振荡培养6 d。菌丝于60 ℃烘干,称重,计算得到菌丝生物量(g/100 mL)。提取并测定菌丝中麦角甾醇含量(mg/g),确定 MeJA 的最佳添加浓度。以MeJA浓度为0的培养基作对照,每个处理三个平行。
(2)SA添加浓度筛选
100 mL液体CYM培养基中分别加入不同体积无菌SA溶液使其终浓度分别为1.00、10.00、100.00、500.00、1000.00 μmol/L,以不含水杨酸的空白培养基作对照,每个处理三个平行。接种及培养条件同上。提取、测定菌丝中麦角甾醇含量,确定SA最佳添加浓度。
1.3.2 添加时间确定
分别在接种的0、2、4、6、8、10 d加入最佳浓度MeJA或SA,于25 ℃,120 r/min振荡培养,并加入空白对照,继续培养至第12 d收获菌丝体,测定生物量和麦角甾醇含量,筛选最佳添加时间。每个处理三个平行。
1.3.3 诱导时长确定
在最佳添加时间(第n d)时加入最佳浓度的MeJA或SA,于25 ℃,120 r/min振荡培养,后分别在第n d、(n+1) d、(n+2) d、(n+3) d、(n+4) d、(n+5) d 收获菌丝,测定生物量和麦角甾醇含量。每个处理三个平行。
1.4 基因表达分析
参考检测获得的 JSR3转录组数据(NCBI:PRJNA305463)以及酿酒酵母(Saccharomycescerevisiae)[14]中报道麦角甾醇合成途径,设计基因表达定量分析引物(表 1)。在优化的条件下分别添加MeJA和SA,收集0~60 h液体培养菌丝液氮速冻后提取总RNA,经反转录后用于定量PCR扩增[15]。扩增体系25.00 μL,反应程序为95 ℃,10 min;95 ℃,15 s→60 ℃,60 s,循环40次;60 ℃→95 ℃,+1 ℃/30 s。以姬松茸内源gpd基因为内参,用2−ΔΔCT法计算基因的相对表达量[16]。
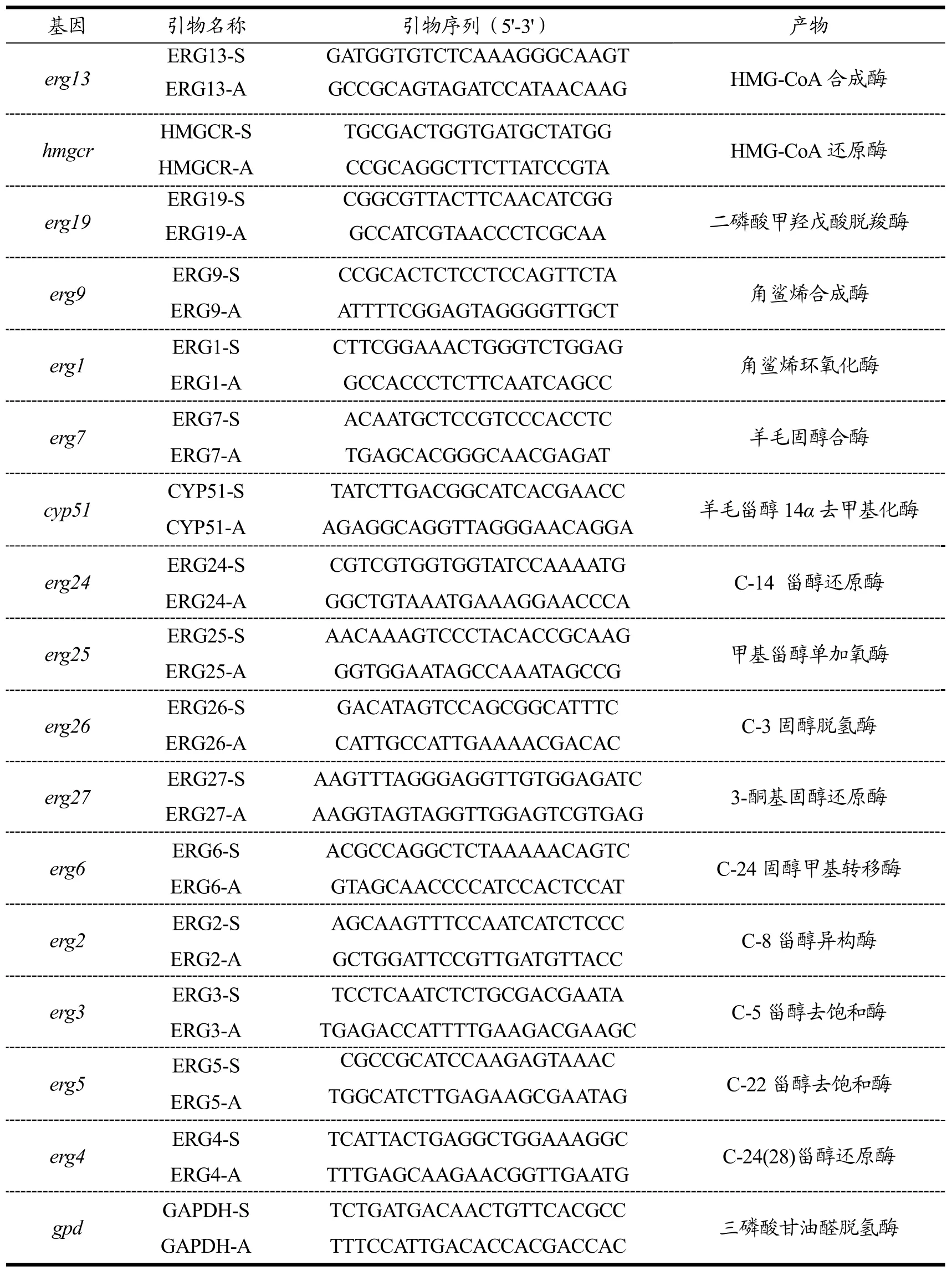
表1 麦角甾醇合成途径中基因表达定量分析引物Table 1 Primers for RT-PCR analysis of genes involving in ergosterol synthesis
1.5 麦角甾醇含量测定
1.5.1 麦角甾醇提取
姬松茸菌丝中麦角甾醇采用碱-醇法提取[17]。称取1.00 g样品溶于32.00 mL碱-醇溶液(KOH 25.00 g,蒸馏水40.00 mL,乙醇定容到100.00 mL),40 ℃下200 W超声20 min,6000 r/min离心10 min,上清液用正己烷萃取2次,收集下层液体水洗到中性,40 ℃旋转蒸干后加入20.00 mL无水乙醇,超声溶解后定容到25.00 mL。
1.5.2 麦角甾醇含量测定
麦角甾醇标准品稀释到 0.02~0.08 mg/mL,采用Inertsil®ODS-SP(5 μm,4.6 mm×250 mm)C18色谱柱,柱温30 ℃,流动相为V(流动相甲醇):V(水)=98:2,流速1.00 mL/min,检测波长282 nm,进样体积10.00 μL。以峰面积Y为纵坐标,标准品进样量X(mg/mL)为横坐标,绘制标准曲线(Y=1×107X+11493,R2=0.9981)。样品稀释或不稀释经0.22 μm滤膜过滤至进样瓶,10.00 μL进样,以峰面积外标法定量。
1.6 统计分析
试验数据利用单因素方差分析(one-way ANOVA test)进行差异显著性分析,以p<0.05判断差异显著性程度。采用Origin 8.0软件进行数据分析作图。利用Hemi软件绘制基因表达量热图。
2 结果与讨论
2.1 外源诱导物添加量筛选
有报道指出,MeJA和SA的浓度会影响细胞的生长和次级代谢产物的产量[9,11-13],因此本研究分别对MeJA和SA诱导姬松茸麦角甾醇合成的最佳浓度进行了筛选。从图1中可以看出,虽然MeJA和SA可以诱导姬松茸麦角甾醇的合成,但麦角甾醇的合成量并不会随着诱导物浓度的增加而持续增加。图1a中,姬松茸菌丝中麦角甾醇的含量随着培养基中MeJA浓度的提高呈现出先增加后减少的趋势,在浓度为70.00 μmol/L时,菌丝中麦角甾醇含量最高,为1.35 mg/g,其含量是对照组(MeJA浓度为0)的1.52倍。但在MeJA不同浓度条件下,菌丝生物量并未表现出与麦角甾醇含量相同的趋势。且与麦角甾醇含量趋势相反,菌丝的生物量在MeJA浓度为70.00 μmol/L时最低,为1.17 g/100 mL,而在不添加MeJA时生物量最高,为1.43 g/100 mL。这一现象与其他物种中报道一致,原因可能是MeJA抑制初级代谢,而激活次级代谢,且MeJA对细胞生长的抑制可能有利于次生代谢物的合成[9,18]。
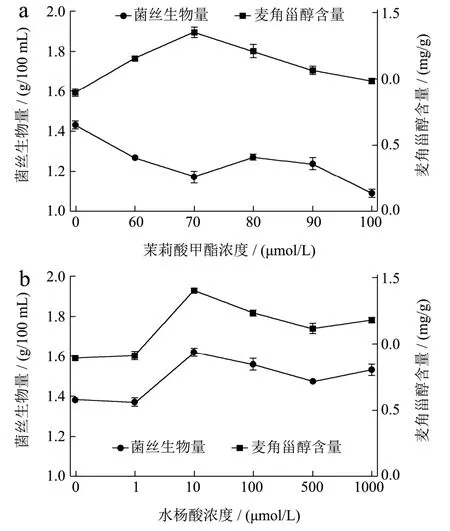
图1 茉莉酸甲酯(a)和水杨酸(b)添加浓度对姬松茸菌丝生物量和麦角甾醇含量的影响Fig.1 Effects of concentrations of MeJA and SA on the mycelial biomass and ergosterol content in Agaricus blazei
与MeJA不同,SA浓度对姬松茸菌丝生物量和麦角甾醇含量的影响具有相同的趋势(图 1b)。虽然生物量和麦角甾醇含量不与 SA浓度成正比,但在 SA浓度为10.00~1000.00 μmol/L时,所有测试浓度均促进了菌丝生物量和代谢产物麦角甾醇的合成,这一结果与 SA 对桑黄(P. igniarius)[12]、猴头菇(H. erinaceus)[13]、灵芝(G. lucidum)[19]中黄酮、麦角甾醇、灵芝酸等次级代谢产物的影响报道一致。在SA浓度为10.00 μmol/L时,姬松茸菌丝生物量和其中麦角甾醇含量达到最大值,分别为1.62 g/100 mL和1.40 mg/g,比对照组(SA浓度为0)分别提高了17.39%和57.30%,表明SA可增加姬松茸中麦角甾醇含量[12,19]。
与本文确定的MeJA和SA对麦角甾醇的最佳诱导浓度(70.00 μmol/L和10.00 μmol/L)不同,猴头菇中报道的MeJA和SA对麦角甾醇的最佳诱导浓度为50.00 μmol/L 和 100.00 μmol/L[13],分析原因可能是不同食用菌菌种对这2种诱导物的耐受性存在差异。
2.2 外源诱导物添加时间
为同时提高麦角甾醇的含量以及菌丝生物量,对外源诱导物的添加时间进行了优化,结果如图2所示。图2a中,培养至第6 d时加入70.00 μmol/L MeJA,培养结束收获的菌丝量达到最大值,为 1.92 g/100 mL,是空白对照组(1.40 g/100 mL)的1.37倍。麦角甾醇的含量随着添加时间的推后,总体呈现先抑制后促进再抑制的趋势,在第8 d添加MeJA时,麦角甾醇含量达到最大值,为2.52 mg/g,是对照组(0.89 mg/g)的2.83倍。而在第4 d加入MeJA,菌丝的生物量和麦角甾醇含量均最小,在第4 d达到最小峰值,推测原因可能是菌丝在刚刚度过“适者生存”期向指数生长期转变时,加入的MeJA后又一次引起“适者生存”现象[20],抑制了菌丝的初级和次级代谢,从而导致菌丝生物量和麦角甾醇含量值最低。考虑到菌丝生物量和麦角甾醇出现最大值的MeJA添加时间不一致,我们以培养终点所获得菌丝中麦角甾醇总量为参考指标,确定第8 d为MeJA的最佳添加时间点,此时获得的麦角甾醇总量最高,为4.56 mg/100 mL。

图2 茉莉酸甲酯(a)和水杨酸(b)添加时间对姬松茸菌丝生物量和麦角甾醇含量的影响Fig.2 Effects of adding time of MeJA and SA on the mycelial biomass and ergosterol content in Agaricus blazei
由图2b可以看出,在12 d的培养周期内,不同时间点加入 SA,菌丝的生物量和麦角甾醇含量表现出相同的趋势。在培养至第6 d加入10.00 μmol/L的SA时最终收获的菌丝量和其中麦角甾醇的含量均达到最大值,分别为2.02 g/100 mL和2.73 mg/g,分别为空白对照组的1.44和3.07倍。根据以上结果,可以看出SA比MeJA具有对姬松茸菌丝量和麦角甾醇具有更好的诱导效果,推测可能是因为二者的诱导机制或作用底物不同[21,22]。
2.3 外源诱导物诱导时长
Shi等[12]研究指出,高浓度或长时间的诱导物处理会造成一个环境压力,从而影响菌丝的生长并引起代谢的改变。因此本研究在已确定的添加浓度和添加时间条件下,分别继续对菌丝进行5 d的培养,每天取样以确定MeJA和SA最佳诱导时长,以提高菌丝生物量和麦角甾醇含量。
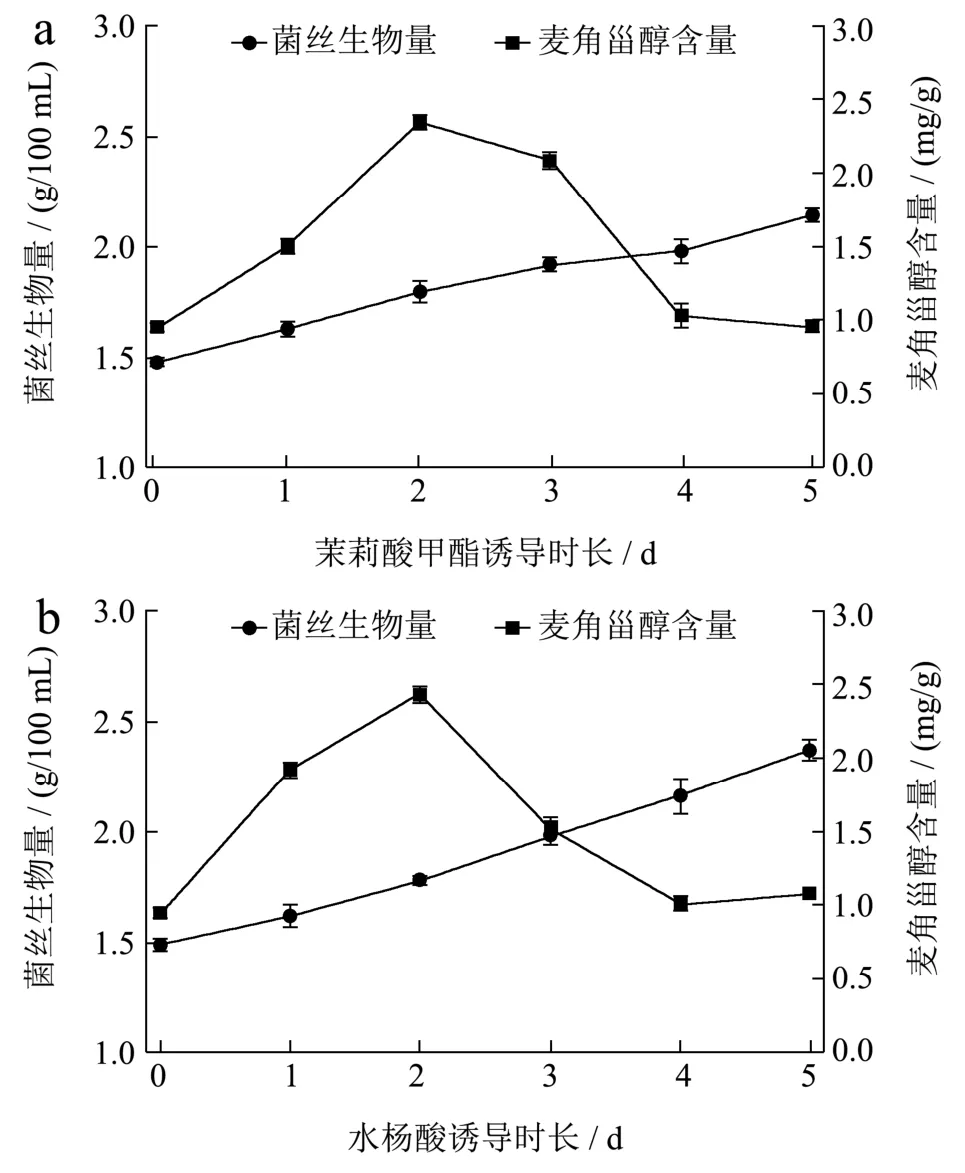
图3 茉莉酸甲酯(a)和水杨酸(b)诱导时长对姬松茸菌丝生物量和麦角甾醇含量的影响Fig.3 Effects of duration of MeJA and SA induction on the mycelial biomass and ergosterol content in Agaricus blazei
由图3可以看出,在加入MeJA和SA后,随着培养时间的延长菌丝生物量基本上呈现出增加的趋势,随着培养时间的延长,菌丝生物量缓慢增加。此外,菌丝中麦角甾醇含量均在添加MeJA和SA后的第2 d达到最大值,分别为2.35 mg/g和2.43 mg/g,比对照组(0 d,0.95 mg/g)分别增加了 147.37%和155.79%。此时,虽然菌丝生物量比较低,但其中麦角甾醇总量均达到最高值,分别为4.24 mg/100 mL和4.33 mg/100 mL。而且,该诱导条件下获得的麦角甾醇含量已超过姬松茸子实体中的含量(1.00~2.00 mg/g)[7],比较姬松茸子实体栽培周期长以及所耗费的人力、物力和财力,本研究所获得的麦角甾醇具有时间短、成本低、产量高等优势,尤其是以SA为诱导物时,培养时间更短而产量更高,因此可以考虑用培养所得菌丝体来代替子实体开展相关研究。
2.4 基因表达分析
Hu等[14]指出麦角甾醇的生物合成途径可分为三个模块,即戊酸合成、焦磷酸法尼酯合成和麦角甾醇合成。合成涉及至少23步反应,25个结构基因[23],这些基因中既有必需基因,也有非必需基因[24]。Ren[11]和Shi[12]等研究指出,MeJA和SA可通过调控参与目标产物合成的基因转录水平或改变次级代谢途径中某些酶的活性来提高目标产物的合成。因此,为探明MeJA和SA诱导姬松茸麦角甾醇合成的机制,本文对添加诱导物前后参与麦角甾醇合成相关酶的编码基因(16个,表1)转录水平进行了定量。图4a和4b分别显示在姬松茸菌丝培养到第8 d加入70.00 μmol/L MeJA和第6 d加入10.00 μmol/L SA后每隔12 h菌丝中参与麦角甾醇合成基因表达情况。

图4 茉莉酸甲酯(a)和水杨酸(b)诱导对姬松茸中麦角甾醇合成相关基因转录水平影响Fig.4 The results of transcription analysis of the genes involving in ergosterol biosynthesis of Agaricus blazeiin response to MeJA(a) and SA (b)
从图4中可以看出,在添加MeJA和SA后,姬松茸菌丝中erg1,erg7,cyp51(erg11)基因的表达量基本都表现为上调,这一结果可能与它们是麦角甾醇合成前期的必需基因有关[14]。此外,erg27的表达量虽也有所提高,但在不同时间点其表达量会出现波动,这可能是其编码的3-酮基固醇还原酶与erg7基因编码的羊毛固醇合酶存在相互调控作用所导致的[25]。
除以上3个基因转录水平明显提高外,在添加SA诱导的中后期,erg6基因表达量出现上调,且随着诱导时间的延长erg4基因的表达量逐渐增加。有报道指出,在酿酒酵母中erg6基因和erg4基因的高表达会增加酿酒酵母中麦角甾醇的合成,预示着它们在麦角甾醇合成中具有重要调控作用[26,27]。据此推测erg6和erg4基因在SA诱导的麦角甾醇含量提高中起到调控作用。但在MeJA诱导的菌丝中未检测到erg6和erg4这2个基因转录水平明显的改变,推测可能是MeJA和SA诱导麦角甾醇合成的机制存在差异。
hmgcr基因编码的HMG-CoA还酶是甾醇生物合成途径中第一个报道的限速步骤,高表达HMG-CoA还原酶可以促进中间体角鲨烯的积累[23],但在 MeJA和SA诱导下(图4),姬松茸菌丝体中hmgcr基因的表达量始终处于一个较低的水平,推测可能是因为较高的麦角甾醇引起的反馈调控[14]。而erg13、erg19、erg5基因也因其表达量过低而未检测到,推测可能跟蔡鹏丽等报道的结果一致,这些基因的表达量与麦角甾醇含量负相关[28],较低的表达量更有利于麦角甾醇的合成。
虽然基因表达量不能完全代表其编码酶的活性,但综上可以推测MeJA和SA诱导麦角甾醇合成主要是通过直接或间接的作用启动麦角甾醇合成上游关键酶基因的表达来提高中间代谢产物的量,并通过中间代谢物与下游相关酶的协同、抑制等互作来实现的麦角甾醇合成的调控。
3 结论
外源添加MeJA和SA可以诱导姬松茸菌丝体中麦角甾醇合成。经过试验获得二者最佳的诱导条件分别是 25 ℃下液体培养的菌丝在第 8 d加入 70.00 μmol/L的MeJA和第6 d加入10.00 μmol/L的SA,并继续培养2 d。获得的菌丝中麦角甾醇含量分别为2.35 mg/g和2.43 mg/g,比对照组分别增加了147.37%和155.79%,并且高于姬松茸子实体中麦角甾醇含量,因此外源诱导,尤其是SA诱导获得菌丝体可替代子实体开展相关研究。通过荧光定量PCR对参与麦角甾醇合成基因表达量进行分析后推测MeJA和SA诱导麦角甾醇合成的机制可能是通过直接或间接的作用启动麦角甾醇合成上游关键酶基因的表达,并通过参与麦角甾醇合成代谢相关酶的相互作用来实现的。
猜你喜欢
现代临床医学(2024年1期)2024-01-30 12:40:22
香格里拉(2023年1期)2024-01-04 02:48:20
食品安全导刊(2022年32期)2022-12-07 05:36:08
中国典型病例大全(2022年10期)2022-05-10 21:24:29
世界最新医学信息文摘(2021年12期)2021-06-09 08:36:46
今日农业(2020年23期)2020-12-15 03:48:26
阿来研究(2018年1期)2018-07-10 09:39:08
知识经济·中国直销(2018年6期)2018-06-29 07:56:26
中国当代医药(2015年36期)2015-03-11 20:03:28
中国粮油学报(2014年8期)2014-02-06 01:34:23
