
萱草糖转运蛋白HfSWEET2a的克隆及低温胁迫下的表达分析
2021-01-05 00:35黄东梅肖海涛张志国秦巧平
应用技术学报 2020年4期
黄东梅,肖海涛,张志国,白 露,秦巧平
(上海应用技术大学 生态技术与工程学院,上海 201418)
糖类物质不仅作为植物体内能源物质供植物生长发育,还参与植物体内的多种信号转导途径[1]。SWEET是一类新发现的具有7个跨膜结构域的糖转运蛋白,该蛋白可作为糖转运载体参与糖的运输[2]。SWEET蛋白有着双向转运的功能,可调节葡萄糖跨膜吸收[3],其广泛存在于原核生物、植物、人类及其他动物中[4]。目前SWEET蛋白在多种作物,如水稻(Oryzasativa)[3]、葡萄(Vitisvinifera)[5]、拟南芥(Arabidopsisthaliana)[6]、高粱(Sorghumbicolor)[7]、大豆(Glycinemax)[8]、番茄(Solanumlycopersicum)[9]等均有相关报道。研究表明,大多数物种SWEET基因家族主要分成4个分支[10],各分支成员之间互相作用共同促进糖的高效运输和细胞生命活动,且对植物种子萌发、胚胎发育、花蜜产生以及花粉发育也起着十分重要的作用[11-12]。
低温胁迫是植物生长过程中经常会遭遇的一种灾害,它不仅影响着植物的生长和产量,而且容易引起植物细胞发生相应的生理变化,甚至有致使植株死亡的可能性[13]。但是也有部分植物在低温胁迫下可以通过对自身生理和生化的相应调整,提高对低温的耐受力,从而达到减轻甚至消除低温胁迫带来的伤害[14]。 由于糖的外排影响着糖的分布变化,植物中SWEET基因的调控对生物和非生物胁迫有着多种影响[15]。研究表明,SWEET基因参与调控植物花蜜分泌[16]、花粉发育[17]、赤霉素的反应[12]、衰老[18]、逆境胁迫[19]、种子和果实发育[20]等细胞生命活动。SWEETs以基因家族形式存在于不同植物中,水稻中鉴定了21个成员[3],大豆中52个[8],拟南芥中17个[6]。Xie等[21]通过对荔枝SWEET基因家族的全基因组鉴定和表达分析发现LcSWEET2a/3b参与了早期种子发育。马铃薯StSWEET2a在糖胁迫后显著上调[22],在高温胁迫下水稻叶片中的OsSWEET2a表达量有着明显的变化[23],枇杷EjSWEET2a-1在8小时盐处理后表达量上调显著[24],以上均表明SWEET2a基因参与了非生物胁迫响应。因此,研究SWEET2a基因应对低温胁迫的响应有着重要的意义。
萱草是深受大众喜爱的花卉之一,‘阿斯隆’是萱草中抗寒性较好的品种,有着较高的利用价值。然而,目前尚未有萱草SWEET基因及其表达调控的有关报道。本研究采用同源克隆的方法从萱草‘阿斯隆’叶片中克隆出SWEET2a基因,利用生物信息学分析该基因的序列特征,同时研究其在低温胁迫处理下的表达情况,旨在为萱草SWEET基因的研究与应用提供一定的参考价值,同时为萱草SWEET基因参与低温胁迫反应的进一步研究奠定基础。
1 材料与方法
1.1 试验材料
萱草‘阿斯隆’(Hemerocallisfulva)植株栽植于上海应用技术大学植物园。将萱草栽植于富含腐殖质的湿润土壤中,置于阳光充足、通风良好的地方培养。5~7天左右浇水一次,正常施肥,根据具体情况进行相应的除草、换盆换土等养护管理。采集萱草‘阿斯隆’长势良好且一致的叶片,用蒸馏水洗净晾干,液氮速冻并于-80 ℃下储存用于后续试验。
1.2 RNA的提取及cDNA的合成
使用改良的CTAB法[25]提取萱草叶片RNA,采用超微量分光光度法和凝胶电泳法检测RNA的浓度、纯度和完整性,-80 ℃保存备用。检测合格的RNA采用M-MuLV 第一链 cDNA 合成试剂盒进行第一链cDNA的合成(上海生工),-20 ℃保存备用。
1.3 基因克隆
以萱草‘阿斯隆’叶片的cDNA为模板,根据转录组测序结果筛选到的SWEET2a基因序列在两端设计引物(见表1),采用PCR技术扩增目的片段并克隆到pUCm-T Vector,进行测序验证得到全长cDNA序列。实验所需引物序列见表1,引物及测序委托生工生物工程(上海)有限公司完成。
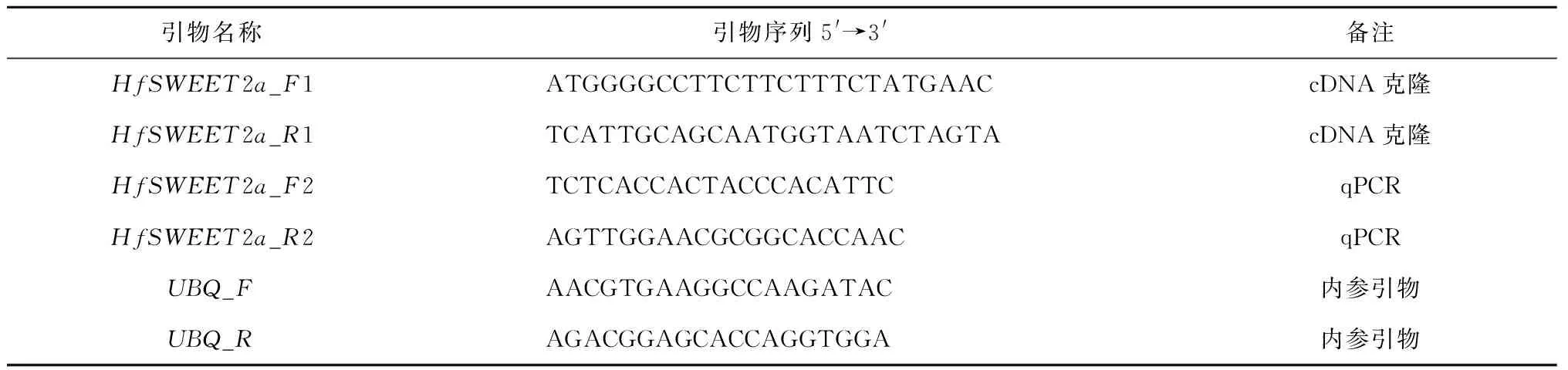
表1 实验所需引物序列Tab.1 Sequence of primers used in this experiment
1.4 基因的生物信息学分析
利用ExPASY(https://web.expasy.org/translate/)网站对SWEET2a核苷酸序列进行翻译,用ProtParam(http://web.expasy.org/protparam)网站进行蛋白质理化性质分析,用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)网站进行亚细胞定位预测,蛋白质跨膜结构分析采用TMHMM在线工具(http://www.cbs.dtu.dk/services/TMHMM/)进行。以HfSWEET2a蛋白序列在NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)网站搜索同源蛋白序列,下载ONK69846.1,芦笋asparagus;XP_008775459.1,海枣date;XP_020701878.1,铁皮石斛dendrobe;THU64444.1,野蕉banana;XP_026394894.1,罂粟opium;OVA00198.1,博落回macleaya;XP_004968893.1,小米millet;XP_021313692.1高粱sorghum;NP_001146103.1,玉米maize;OAY69557.1,菠萝pineapple;XP_020595974.1,蝴蝶兰orchid;MQL94069.1,芋头taro;RWR97356.1,沉水樟cinnamomun;XP_031490470.1,蓝星睡莲lotus;XP_012848541.1,猴面花guttata;XP_024020545.1,川桑mulberry;XP_011623118.1,无油樟amborella;TXG47757.1,漾濞槭acer;XP_017234459.1,黄胡萝卜carrot;XP_006467563.1,橙orange;XP_022715681.1,榴梿durian;QHT64202.1,荔枝Litchi;KAF3666069.1,辣椒pepper;AMQ35580.1,马铃薯potato;QIE48602.1,茉莉花jasmine;XP_028118815.1,茶camellia;XP_007025233.2,可可树cacao;XP_020553914.1,胡麻flax等28种SWEET2a蛋白序列进行后续分析。利用Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)和jalview2.10.2软件进行多序列比对,再用MEGA7.0软件输出基于neighhor-joining法构建的系统进化树,bootstrap值设置为500次重复。用MEME(http://meme-suite.org/tools/meme)进行motif比对,再用NCBI的CD-Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)进行保守结构域查询。
1.5 低温处理及基因定量表达分析
为进一步了解基因对低温胁迫的响应,选长势一致的萱草幼苗移入低温人工气候箱中,分别置于15 ℃、10 ℃、5 ℃、0 ℃下处理,24 h后进行取样,取样部位为完全展开的功能叶片,设置3次生物学重复。利用Primer5(https://sg.idtdna.com/pages/tools/primerquest)网站,根据HfSWEET2a基因cDNA序列设计定量表达分析所用引物HfSWEET2a_F2/HfSWEET2a_R2,内参为UBQ(见表1)。提取叶片RNA、反转录获得cDNA后,进行qPCR,反应条件为95 ℃预变性5 min;95 ℃变性10 s,60 ℃退火30 s,循环40次;融解曲线采集程序为95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。采用2-ΔΔCt法计算基因相对表达量,同时计算其标准差,再用Excel和SPSS22.0对数据进行处理。
2 结果与分析
2.1 萱草SWEET2a基因的克隆及序列分析
以萱草‘阿斯隆’叶片的cDNA为模板,通过PCR技术克隆得到长度为 1 045 bp的核苷酸序列,命名为HfSWEET2a,该基因已经提交GenBank,登录号为MT379659。序列分析显示,HfSWEET2a含有690 bp的完整开放式阅读框,编码229个氨基酸(图1)。蛋白质理化性质分析表明,HfSWEET2a编码的蛋白分子量25.531 kDa,等电点9.16,定位在细胞膜。蛋白质跨膜结构分析显示,HfSWEET2a含有典型的7个跨膜结构域,其位置分别为:10-32,45-67,71-93,105-127,137-159,166-188,193-215(见图2)。

下划线表示PCR引物序列,黑色标出字母分别表示起始密码子和终止密码子灰色部分代表两个保守结构域,框出部分代表7个跨膜结构域(TM1-7)图1 萱草SWEET2a基因的核苷酸序列及编码氨基酸序列Fig.1 Nucleotide sequence and encoding amino acid sequence of HfSWEET2a

图2 HfSWEET2a蛋白的跨膜结构预测Fig.2 Transmembrane probabilities for HfSWEET2a
2.2 萱草HfSWEET2a基因编码蛋白与其他植物进化关系分析
BLASTX比对结果表明,HfSWEET2a的蛋白序列与芦笋、菠萝、野蕉等SWEET2a蛋白序列相似性较高,分别为75.58%、74.6%、73.09%。将HfSWEET2a蛋白与芦笋ONK69846.1等10条蛋白质序列进行序列比对,结果显示HfSWEET2a与这些蛋白的保守结构域高度相似。motif比对结果显示,它们都有3个相同的motif,且均为PQ-loop家族(见图3中的PQ-loop 1、PQ-loop 2和PQ-loop 3)。
系统进化分析结果显示单子叶和双子叶SWEET2a蛋白分别聚为两类,HfSWEET2a与芦笋的系统进化关系最近,聚在同一分支,其次为菠萝、野蕉和海枣,与双子叶植物荔枝、胡麻以及茉莉花等的亲缘关系较远(见图4)。

图3 萱草HfSWEET2a编码蛋白与其他10种植物SWEET2a蛋白的motif比对Fig.3 The motif comparison of HfSWEET2a and SWEET2a proteins from other 10 plants
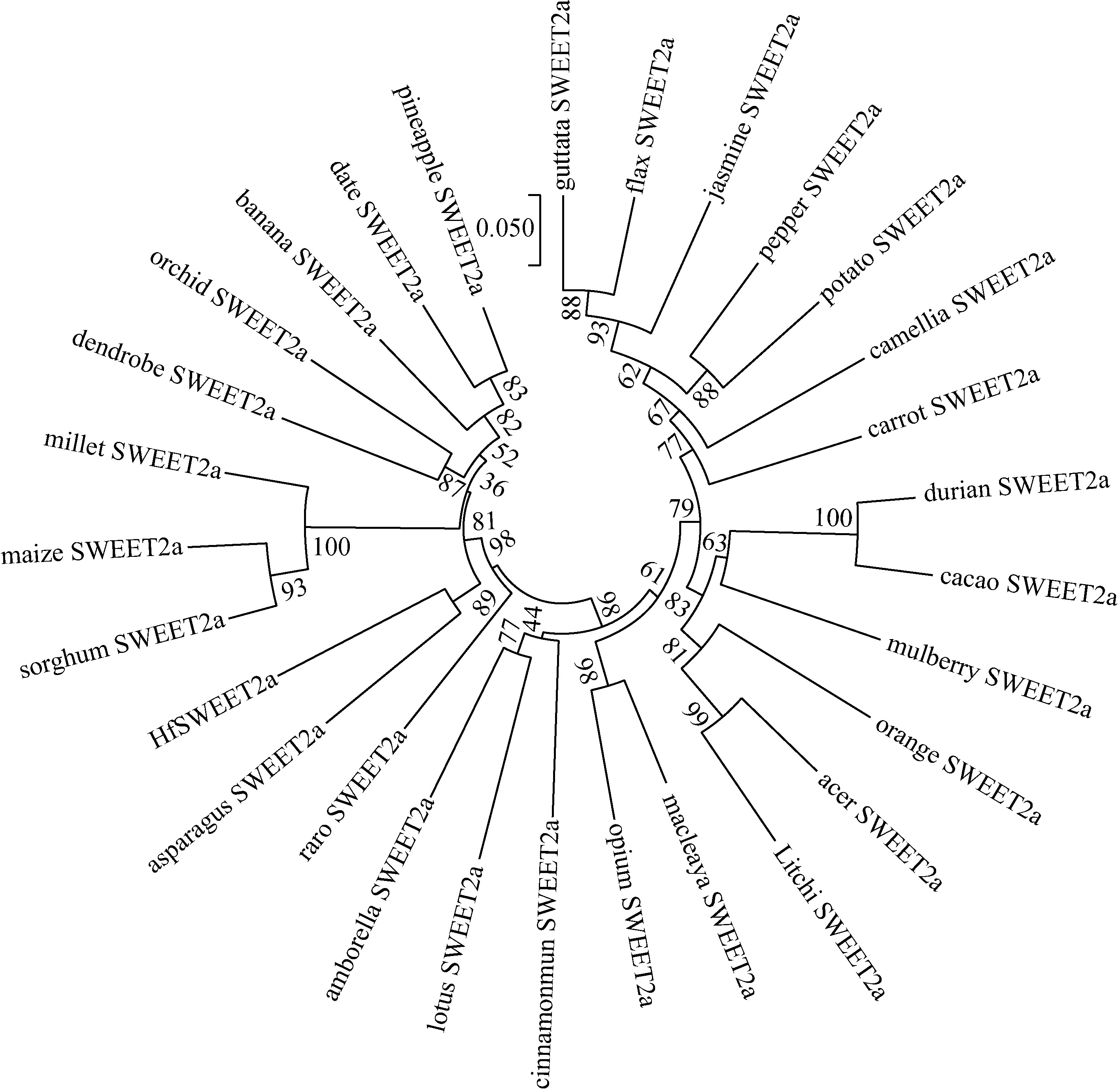
图4 萱草HfSWEET2a与其他28种植物SWEET2a蛋白的系统进化分析Fig.4 The phylogenetic analysis HfSWEET2a and SWEET2a proteins from other 28 plants
2.3 低温对萱草HfSWEET2a基因表达的影响
基因定量表达分析显示,在不同温度处理下,HfSWEET2a的表达呈现出不同程度的变化。HfSWEET2a基因在0 ℃下表达量最高,0 ℃相对表达量是15 ℃下的3.3倍、10 ℃下的1.7倍、5 ℃下的3.7倍(见图5)。统计分析显示,5 ℃和15 ℃下的HfSWEET2a基因表达差异不显著,其他温度下的基因表达均达到显著差异水平(P<0.05)。
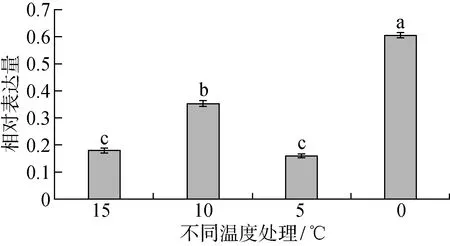
柱形表示平均值±标准误,相同字母表示差异不显著图5 不同温度处理后HfSWEET2a基因的表达变化Fig.5 The expression of HfSWEET2a under different temperature
3 讨论
糖是植物光合作用的主要产物,也是植物体内的主要能源物质[1]。SWEET蛋白广泛存在于各种植物中,参与植物生长发育和逆境响应。目前已有大豆[8]、拟南芥[6]、小麦(Triticumaestivum)[26]、陆地棉(Gossypiumhirsutum)[27]等植物上均有关于SWEET蛋白的研究报道,但萱草作为我国传统花卉之一,目前未见有关SWEET基因的相关报道。本研究通过PCR技术从萱草‘阿斯隆’叶片中克隆出HfSWEET2a的全长cDNA序列,同源序列比对结果表明,HfSWEET2a基因所编码的蛋白质序列与芦笋、菠萝以及海枣等植物SWEET蛋白质序列同源性较高,具有高度相似的保守结构域,且具有均为PQ-loop家族的3个相同的motif。
SWEET家族基因一般都具有典型的6~7个跨膜结构域,菠萝中39个SWEET蛋白中大部分包含7个ɑ-螺旋的跨膜结构域[10],水稻中有18条OsSWEET蛋白具有6-7个跨膜结构域[28],雷蒙德氏棉SWEET基因家族跨膜结构域预测结果显示有22个SWEET蛋白含有7个跨膜结构域,7个SWEET蛋白含有6个跨膜结构域[29]。这些跨膜结构域使其具有保守的跨膜糖转运功能以及介导植物对疾病的感病功能[30],本研究分离到的HfSWEET2a与其他植物SWEET蛋白相似,具有7个保守的跨膜结构域,故HfSWEET2a基因属于SWEET基因家族,可能参与糖的跨膜运输。
低温逆境胁迫是植物中常见的一种非生物胁迫,在低温逆境胁迫下,会有很多结构及生理生化方面的变化,其中可溶性糖含量变化是重要指标之一[31]。可溶性糖是植物在低温逆境下的呼吸作用底物,对保护物质起渗透作用,可增强植物的耐寒性[32-33]。当遭遇低温胁迫环境时,为减少逆境胁迫带来的伤害,植物体内糖类含量会有明显的增长[34-35]。SWEET蛋白作为糖转运载体参与糖的运输、分配和贮藏,被运输到植物体内不同组织、细胞器中,参与植物生长发育的重要生理过程[36],不仅可以响应生物胁迫,而且也是非生物胁迫响应的重要蛋白,SWEETs可以通过调控糖的运输和分配来参与非生物胁迫的响应[37]。
许多植物基因低温响应的顺式元件LTR位于SWEET基因的启动子区域[38],SWEET蛋白的中断对拟南芥的耐寒性有较大的负面影响,耐寒力远不如正常植株[39],而AtSWEET16过量表达植物却比平常表现出更好的抗冻性[40]。CsSWEET16也被证明在改善拟南芥的耐寒性方面了发挥了重要作用[19]。在本研究中,萱草叶片在不同低温情况处理,HfSWEET2a基因表达发生了不同程度的变化,这与茶树、番茄、甘蓝等研究中SWEET基因的表达较为一致。茶树在冷训化过程中,不同SWEET基因的表达量有着不同的变化,CsSWEET2、CsSWEET3和CsSWEET16的表达受到了广泛的抑制,而CsSWEET17和CsSWEET1的表达量却有着显著地提高[41]。番茄中9种SWEET基因在低温处理下叶片中的表达量上调了好几倍,而在根中则显著下调[42]。甘蓝中BoSWEET16a和BoSWEET17的表达在低温胁迫后迅速下降,并在12 h至48 h保持低水平状态[43]。本研究初步表明,HfSWEET2a基因可能参与了低温胁迫应答反应并发挥着重要作用,具体作用机制有待进一步研究。
猜你喜欢
中华实验眼科杂志(2022年8期)2022-11-15
湖北农业科学(2022年11期)2022-07-18
紫禁城(2020年5期)2021-01-07
医药前沿(2020年28期)2020-12-02
生物信息学(2020年1期)2020-05-16
心肺血管病杂志(2019年1期)2019-04-22
教学考试(高考生物)(2017年4期)2017-12-13
天津科技大学学报(2016年1期)2016-02-28
海峡姐妹(2016年7期)2016-02-27
人间(2015年19期)2016-01-04
