
典型模式微生物基因表达精细调控工具的研究进展
2020-11-18 07:14田荣臻刘延峰李江华刘龙堵国成
合成生物学 2020年4期
田荣臻,刘延峰,李江华,刘龙,堵国成
(1 江南大学糖化学与生物技术教育部重点实验室,江苏 无锡 214122; 2 江南大学工业生物技术教育部重点实验室,江苏 无锡 214122)
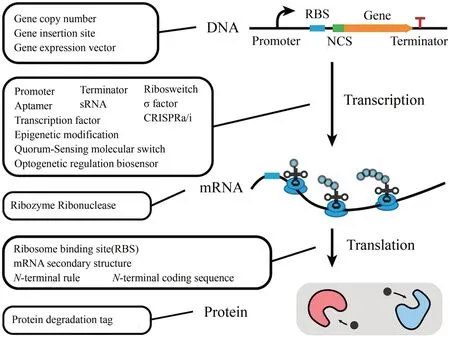
随着基因编辑、高通量基因测序技术的飞速发展,典型模型微生物大肠杆菌(革兰氏阴性细菌)、枯草芽孢杆菌(革兰氏阳性细菌)和酿酒酵母(真核微生物)中新型基因表达调控工具的研究取得了显著进展,进一步推动了合成生物学和代谢工程的发展。为了满足合成生物学研究中对基因表达调控工具动态范围和动态模式的需求,研究人员已经设计、构建和测试了一系列用于合成生物学研究中精确和动态控制基因表达的新型工具[1-2]。本文作者首先简要总结了经典的基因表达调控元件,包括启动子、核糖体结合位点(RBS)和终止子等。在此基础上,进一步总结了近年来发展起来的具有更大的动态范围和更广阔的应用领域的新一代基因表达元件,例如基于大数据的合成启动子、N 端编码序列和小转录激活RNA。接下来,总结了可以实现全局基因调控的工具,例如基于CRISPR-dCas9 系统的基因表达调控工具、基于全局转录因子的基因表达调控工具、基于σ因子的基因表达调控工具和基于表观遗传修饰的基因表达调控工具。此外,还讨论了近年来可以响应特定信号的基因表达调控工具的开发,例如光遗传调节系统、群体感应系统、生物传感器系统和蛋白质降解系统等。最后,提出了目前基因表达调控工具发展中仍然需要解决的问题,如响应元件的特异性、不同基因表达调控工具的标准化以及新型基因表达调控元件在工业规模上的应用,并针对新型基因调控工具设计、构建和应用的发展趋势进行了展望。
1 经典基因表达调控元件
细胞内基因表达水平的调控是多层次、复杂的过程,涉及基因转录、翻译及蛋白降解等,通过相应基因表达元件强度而改变基因表达水平是经典基因表达调控元件探索与应用的基本思路,也是实现胞内基因表达最简便有效的方式之一。经典的合成生物学基因表达调控元件,如启动子、RBS、终止子等可以有效调节转录水平与翻译水平,在调控蛋白表达水平、改变胞内代谢流等过程中发挥了重要作用。目前针对典型的模式微生物,如大肠杆菌、枯草芽孢杆菌、酿酒酵母等均已构建了相应的数据库,以提供不同强度和不同类型的基因表达元件。随着系统生物学,特别是转录组学和蛋白组学测定技术的发展,不同强度、不同类型的经典基因表达调控元件库的构建变得更简便和高效,这进一步增强了对基因表达水平的精确调控能力。
1.1 启动子
启动子是合成生物学和代谢工程领域中使用最广泛、最基本的表达元件,也是最简便有效的基因表达调控方法之一。它是一段能够被RNA 聚合酶识别并结合,从而对特定基因实现转录的DNA 序列。目前,对经典模式微生物均构建了包含不同类型和强度启动子的文库。这些启动子大部分是微生物基因组上固有的启动子序列,通过使用易于检测的报告蛋白(如绿色荧光蛋白GFP、半乳糖苷酶等),在特定培养条件下进行定量表征后得到的。例如在枯草芽孢杆菌中,分别以GFP、脂肪酶、角蛋白酶和碱性果胶酶为报告基因,表征了基因组上原始的114 个启动子,构建了包含3种不同类型、适用于不同pH 和温度、表达强度覆盖约70 倍的启动子文库[3]。但是,由于文库构建通常以报告基因表达量间接表征启动子的转录强度,当使用相同的启动子表达其他基因时,可能存在难以预测的差距[4]。同时,传统的启动子文库构建方式存在的耗时耗力、成本高等问题亟待解决。随着系统生物学,特别是转录组学的发展,胞内所有转录得到的mRNA 序列及丰度等数据可以被精确测量,这为启动子文库的丰富提供了理性的参考数据。
在所有表征的启动子中,大部分启动子为组成型启动子,其不需额外的诱导即可使基因在胞内持续表达。除此之外,经典的启动子中也包括少数在生物体内发现的诱导型启动子,其在胞内的表达受到特定条件,如特定化合物的诱导。由于微生物的生存必须适应不断改变的生活环境,细胞内对于特定基因表达的开启或关闭十分重要。其中,研究最透彻且最早开始使用的基因表达调控元件之一便是基于乳糖操纵子的乳糖诱导型启动子[5]。乳糖操纵子有4 个基本的组成部分:调节基因、启动子、操纵基因和结构基因。当细胞处于无乳糖的培养基时,调节基因表达得到的调节蛋白结合于启动子和结构基因间的操纵基因上,使得启动子难以转录其后乳糖代谢相关结构基因。当环境中存在乳糖时,操纵基因上的调节蛋白脱落,使得启动子正常转录其后的结构基因。根据这一原理,设计了乳糖诱导型启动子用于合成生物学领域中对基因表达强度的调控。在实际应用中,由于异丙基-β-D-硫代半乳糖苷(IPTG)同样可使调节蛋白脱落,且由于其更加稳定、不易被细胞降解的特性,常被用于乳糖诱导型启动子的诱导剂以激活启动子功能。这些启动子的开发可以人为对基因表达时机和强度进行干预,因此在合成生物学中具有重要的意义。
1.2 RBS与终止子
在原核生物中,转录得到的mRNA 通过5′端RBS 序列实现核糖体的招募与结合,进一步实现基因的翻译过程。因此,RBS 的强度直接影响了核糖体的结合效率并在一定程度上决定了翻译强度。相比于启动子,RBS 由于序列较短、操作便捷且调控范围较大,同样是经典的微生物基因表达调控元件之一。目前,通过对大量RBS 序列与强度的统计与分析,认为核糖体与RBS 的结合取决于核糖体结合速率、折叠动力学和折叠能量学。基于此,已经有可以在一定程度上辅助设计RBS 与预测RBS 相对强度的计算工具,如RBS Calculator[6-7]。
基因转录过程的终止,需要一段特殊的DNA序列为RNA 聚合酶转录提供终止信号,这段序列通常位于原核生物基因簇最后一个编码基因之后或真核生物所有编码基因之后,称为终止子。终止子的释放效率对于提高转录效率与提高mRNA稳定性十分重要,因此,目前也开发了一系列典型模式微生物的终止子文库。例如,对于枯草芽孢杆菌,基于λ 噬菌体进行了终止子突变文库的构建,得到了具有更高转录终止效率的终止子突变体和利于mRNA 稳定性的终止子突变体,并且这些方法同样适用于其他典型模式微生物[8]。
2 新型基因表达调控元件
随着合成生物学的快速发展,经典的基因表达调控元件在普遍应用的同时,亟待其性能上的提升以满足研究中日益增长的需要。首先,理想的基因表达调控元件应具有调控范围大的特征,而从基因组上鉴定的天然调控元件强度由于受到进化的限制而不足以为合成生物学提供所需要的强度。其次,理想的基因表达调控系统应具有稳定的特征,诱导型启动子应具有基础表达严密的特征,而经典的天然基因表达调控元件在细胞内潜在的调控机制可能干扰基因的稳定表达[9]。此外,由于对中心法则认识的有限,经典基因表达调控元件在基因表达的不同层次上往往存在缺失。因此,为了进一步实现基因表达的精确调控并扩大调控强度的范围,逐渐开发应用了一些新型的基因表达调控元件。
2.1 新型启动子
2.1.1 合成启动子
经典的启动子往往来源于筛选表征得到的天然启动子,而胞内单基因的过量表达往往会损害细胞的生长,不利于宿主在自然选择中的生存,因此经典启动子的强度往往受到很大的限制。在合成生物学研究与代谢工程改造中,天然启动子无法满足对于超高强度启动子的需求。因此,新一代合成启动子文库逐渐被构建和应用。新型启动子文库根据构建方式一般分为3种,即易错PCR(聚合酶链反应)启动子文库、启动子间隔区域饱和突变文库和混合启动子文库[4]。易错PCR 文库中的启动子是在天然启动子的基础上,通过易错PCR 将随机突变引入启动子的关键序列而得到的。其优点是可以将随机突变引入整个启动子区域,从而覆盖尽量多的突变可能性,但是这个过程往往引入大量使启动子失效的突变,使得文库正向率较低。启动子间隔区域的饱和突变方法将随机突变靶向引入到启动子保守序列-35 区与-10 区之间,可以在不影响启动子功能的前提下,获得正向率较高的启动子文库。混合启动子工程是目前从实验结果来看获得超强启动子最高效的方法。其在已知的天然高强度启动子的基础上,将启动子从两个保守区域-35 区、-10 区截断为3 个序列模块,然后将模块自由组装搭配,得到最终的混合启动子文库,通过使用流式细胞仪筛选出强度最高的启动子。通过这种策略,已经在枯草芽孢杆菌中构建得到了强于通用启动子P43约500 倍的超高强度合成启动子[10]。
2.1.2 广谱启动子
在不同的典型模式微生物,如大肠杆菌、枯草芽孢杆菌与酿酒酵母中表达相同的基因以进行合成生物学研究或代谢工程改造和在不同典型模式微生物中对未知DNA 片段或基因簇的鉴定和功能表征是有效的技术手段。但是,目前天然表征的启动子在不同物种中往往是不相容的,特别是在原核微生物和真核微生物之间,这导致在不同宿主中进行相同基因表达时需要重构具有物种特异性启动子的表达框。为了解决此类问题,目前已经开发了具有不同强度并且典型模式微生物间通用的广谱启动子[11]。这些广谱启动子可用于优化不同宿主中的遗传回路或生物合成途径,从而简化或缩短操作过程。
2.2 基于功能RNA 或DNA 的新型基因表达调控元件
2.2.1 核糖开关
核糖开关是某些mRNA 非编码部分内的结构域,其可以响应特定代谢物的结合而改变自身结构来控制基因表达[12-13]。在细菌中,核糖开关由执行配体识别的适体域和调节转录终止或翻译起始的表达平台组成。适体域在不同生物中高度保守,其作为目标代谢物的精确传感器,具有高度的选择性和特异性。而表达平台在序列和结构上具有较大的可变性,其可以将代谢物与适体域的结合或脱离事件,转化为其后基因表达的开启或关闭。同时,核糖开关可以控制ρ因子依赖的转录终止以调控基因的表达。除了可以实现调控基因表达的功能,核糖开关还可以参与mRNA降解的调控[14]。例如,在大肠杆菌中发现的赖氨酸核糖开关,在赖氨酸结合的情况下,表达平台中RBS 序列形成二级结构,阻止其后的基因进行翻译,同时,赖氨酸的结合暴露了表达平台中RNA降解酶RNase E的切割位点,加速了相应mRNA 的降解过程。而在赖氨酸未结合的情况下,RBS 序列暴露,而表达平台中的RNase E结合位点被隐藏,使得其后的基因得以实现翻译[15](图1)。
由于大部分核糖开关发现于原核生物,因此,天然和合成的核糖开关在大肠杆菌和枯草芽孢杆菌中得到了广泛的应用。如在大肠杆菌中,将上述赖氨酸核糖开关与细胞的生长相偶联,即高产赖氨酸可以同时使细胞获得快速生长的特性,经过定向进化,可获得高产赖氨酸的工程大肠杆菌。同时,核糖开关也广泛应用于大肠杆菌中其他氨基酸类和高附加值化合物的高效生物合成,如色氨酸、维生素B12、N-乙酰神经氨酸(NeuAc)和柚皮素等[16-19]。在枯草芽孢杆菌中,通过将glmS核糖开关整合至NeuAc 生物合成途径关键基因的表达框中,NeuAc 合成的分支途径和中心代谢途径强度得以调控,提高了NeuAc 的产量。在真核典型模式微生物酿酒酵母中,由于其翻译元器件与原核生物的明显差别,导致核糖开关在胞内直接应用较为困难。但是,在酵母中开发了一种间接利用核糖开关的策略,即通过利用核糖开关在配体结合时的自切割活性以控制翻译水平,如在酿酒酵母中开发的新霉素开关[20]。
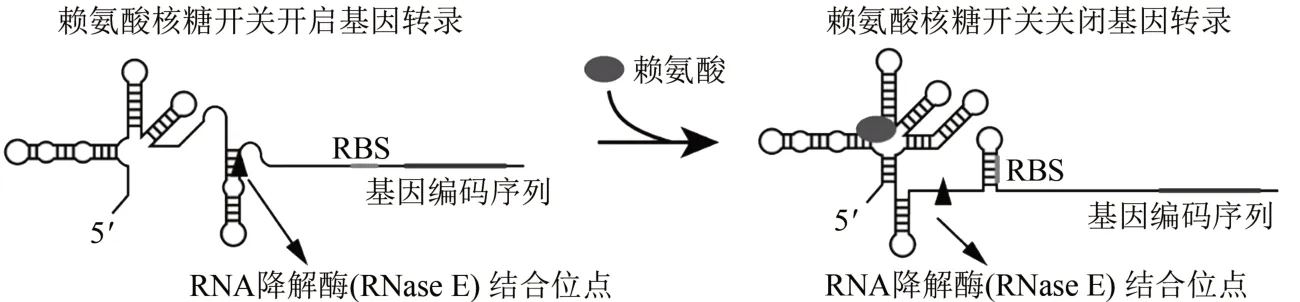
图1 大肠杆菌赖氨酸核糖开关基因表达调控方式Fig.1 Regulation of gene expression based on E.coli lysine riboswitch
2.2.2 sRNA
细菌功能RNA 调控元件的另一类是一种较短的RNA,称为小RNA(small RNA,sRNA)。大部分sRNA通过与特定mRNA直接碱基配对而影响基因的表达,但也有少部分通过直接与蛋白质发生相互作用从而调控蛋白活性[12]。sRNA是胞内转录后调节因子中最丰富的一类,并参与细胞中众多的代谢和应激反应过程。根据与mRNA 配对方式的不同,将sRNA 分为顺式作用与反式作用。此外,RNA 伴侣Hfq 可以通过促进sRNA 碱基与其目标mRNA 的配对,从而充当sRNA 依赖性基因表达的关键介体。在所有的sRNA中,大肠杆菌rpoS基因的5′非翻译区是研究最透彻的序列之一。RpoS蛋白是RNA聚合酶(RNAP)的σS亚基,而σS亚基是细胞响应压力时最普遍的σ因子之一,其调控细胞处于饥饿和不利生存环境时基因的表达[21]。rpoS的表达受到DsrA、RprA和ArcZ这3种sRNA的正调控。在对数生长期间,这些sRNA 的表达水平较低,并且rpoS的翻译受到其mRNA 5′非翻译区中茎环结构的抑制,从而阻止了核糖体RBS结合以抑制基因的翻译过程。在细胞处于稳定期或应激状态时,积累的DsrA、RprA和ArcZ通过与mRNA 5′非翻译区中碱基配对以实现茎环结构的破坏,使RBS序列暴露从而减轻对翻译的抑制。同时,sRNA 可以通过抑制控制ρ因子对基因的提前终止以调控基因的表达,实现细胞对不断变化的生长条件的响应。
在多种sRNA 中,asRNA(antisense RNA,反义RNA)广泛应用于代谢工程中的基因表达调控[22]。其在基因组上的启动子方向与调控目的基因相反,但转录重合的一段DNA 序列,因此,asRNA 与目的mRNA 完全互补。asRNA 通过两种方式进行对基因的调控,一种是通过在基因组上实现反义转录时,可以与正向转录的RNA 聚合酶发生碰撞脱落,从而对正向的基因转录起到一定的抑制作用;另一种是asRNA 与转录得到的目的mRNA 的互补结合对目的基因翻译起到抑制作用。目前,asRNA 调控已经应用于模式微生物中葡萄糖酸、白藜芦醇、柚皮素和4-羟基香豆素等高附加值化合物的生物合成[23-24]。
2.2.3 核酸适配体
核酸适配体是近年来开发的一种由单链DNA或RNA 适配体和蛋白配体组成的新型基因表达调控元件。核酸适配体与核糖开关不同,核糖开关是与mRNA 共转录的RNA 元件,通过自身结构改变来调节基因表达;而核酸适配体可以是单链DNA 或RNA,其通过识别和结合配体(包括蛋白质和小分子)发挥功能[25]。适配体可以通过特异性结合相应的配体来改变其空间构型以进一步调节下游基因的转录或翻译。迄今为止,研究人员已经针对各种靶标(包括蛋白质和小分子)鉴定了数千种DNA 或RNA 适配体[26]。其中,DNA 适配体介导的转录激活依赖于两个配体分子与两个单链DNA适配体的结合,这种结合使得DNA双链被解旋,从而有利于启动子转录的进行[27-28]。
基于适配体动态增强基因表达水平的原理,已经构建了多种基于核酸适配体-配体的新型基因表达元件。例如,在枯草芽孢杆菌中,基于DNA 适配体与凝血酶配体构建并优化了用于上调基因表达水平的基因表达元件,基于RNA 适配体与凝血酶配体构建并优化了用于下调基因表达水平的基因表达元件。在合成2′-岩藻糖基乳糖的工程枯草芽孢杆菌构建过程中,通过使用该上调和下调元件分别调节了外源途径基因的表达和内源乳糖运输抑制基因的表达,使2′-FL 的产量提高27.3 倍,充分表明了该元件的有效性[29]。
2.3 N 端编码序列
精确控制基因表达强度和动态模式对于代谢工程和合成生物学非常重要,特别是对于精细调控代谢途径和优化基因回路。经典的调控基因表达的基因调控元件,包括启动子、终止子、RBS等,虽然针对中心法则的每一环节均提出了相应的调控方式,但是在每个环节仍然存在对基因表达量影响巨大的其他因素。其中目的基因本身的序列,特别是基因N 端编码序列(N-terminal coding sequence,NCS)便是其中重要的影响因素之一,其通过影响核糖体在翻译起始阶段与mRNA 结合和延伸的效率,在翻译水平上强烈影响基因表达,是细菌中的基因表达水平调控的重要机制[30-33]。在近年来的研究中,通过在典型模式微生物中的表征,越来越多的研究证实了N 端编码序列对于基因表达的重要影响,并且提出了潜在的6 种N 端编码序列影响mRNA 翻译水平的机制,包括N 端规则、mRNA 二级结构、热力学和动力学、AT 含量、带电荷数、氨基酸残基数以及特殊氨基酸的丰度[34-39]。
目前,使用工程化的NCS 工具在代谢工程改造中精细调控代谢途径仍然十分具有挑战性。尽管如此,在合成生物学研究和代谢工程的实际应用中,N 端编码序列的重要性正被逐渐重视。例如,在大肠杆菌中,通过对于超过24.4万个合成N端编码序列文库的构建、高通量测序、表征和分析,为N端编码序列的进一步机理研究和代谢工程应用提供了基础[40]。在枯草芽孢杆菌中,目前开发了天然的和合成的N端编码序列文库及N端编码序列的改造方法以精细调控基因的表达。在这个过程中,首先构建和表征了基于系统生物学的N端编码序列文库作为基因表达调控元件并将其分为了4 个类型,即生长偶联型、持续表达型、迟滞表达型和强烈抑制型。然后,根据对于文库序列的统计分析,设计了适用于枯草芽孢杆菌的工程N端编码序列设计规则。最后,天然和合成的N端编码序列作为基因表达调控元件成功应用于NeuAc的生物合成途径中[41]。这些研究表明N 端编码序列可用作典型模式微生物的新型的基因表达调控元件。
3 基因表达的全局调控工具箱
经典的基因表达调控工具虽然可以实现对单基因表达水平的调控,但在合成生物学和代谢工程中的实际应用往往需要对多基因表达调控的配合。这限制了对宿主进行改造的通量,限制了同时探索多个基因的可能,使细胞难以实现全局的最佳表型[2]。同时,细胞内代谢网络往往存在复杂的调控机制,在胞内进行单基因或多基因表达强度的改变或引入异源代谢途径往往造成细胞内代谢的失衡,代谢负荷加剧,进一步损害细胞生长,不利于代谢工程菌株在工业上的大规模应用[42-43]。胞内天然代谢网络的调控机制为解决这些限制提供了方法。当环境条件发生变化或细胞处于不利环境时,胞内代谢网络可以迅速响应环境的变化,开启或增强当前环境条件下利于细胞生存的一系列基因的表达,并抑制对细胞不利基因的表达。这种全局性的基因表达调控使细胞能够适应不同生长环境中的各种营养条件,诱导相应环境条件下合适的细胞过程,如细胞发育、氮代谢和碳代谢等,提高细胞在不同环境下的抗逆性。而这些全局性的基因表达调控系统往往受调控于某些特定的蛋白或RNA,如全局转录调节因子、σ 因子、6S RNA 和DNA 修饰酶等。因此,通过改变相应基因的表达水平,即可实现胞内全局性的基因表达精确调控。此外,近年来由于合成生物学技术的快速发展,也产生了人工的高通量基因表达调控工具——基于CRISPR-dCas9 系统的基因表达调控工具,其可以在不改变目的基因启动子的情况下实现对多基因进行全局调控。同时,使用σ因子进行胞内全局转录调控的策略被称为全局转录机器工程(gTME)[44]。目前,此概念已经拓展至对所有的全局基因表达调控工具的应用策略。
3.1 基于全局转录调节因子的全局基因表达调控工具
全局转录调节因子是一类特定的功能蛋白,其通过与目标基因启动子区域或附近的序列特异性结合,充当基因表达的阻遏物或激活物。其存在使细菌能够整合各种信号,以响应营养物质、活性氧、渗透压等其他环境因素的变化而改变其基因表达程序[45-46]。在原核与真核微生物中,可以通过对全局转录调节因子的表达水平调控或特异性位点突变,实现对某一类功能基因的全局性调控。
目前在3种典型模式微生物中均已实现了全局转录因子对于全局基因表达调控的应用。在大肠杆菌中,通过定向进化,得到了耐受并高产3-羟基丙酸的工程菌株。通过全基因组测序、基因回复突变等表征手段,发现YieP 全球转录调节因子的突变增强了大肠杆菌对3-羟基丙酸的耐受性[47]。在枯草芽孢杆菌中,开发了基于全局转录因子CodY 和CcpA 的碳氮代谢网络调控方法,通过对两种全局转录因子突变文库的构建与表征,成功实现了对于碳源和氮源吸收和利用的全局性调节,大大提高了β-半乳糖苷酶、木聚糖酶和肽酶的产量[48-49]。在酿酒酵母中,通过对转录调节因子SPT3 的随机突变,成功提高了其铅耐受性[50];通过对转录调节因子Pdrlp 的突变,可以实现其对有机溶剂的耐受性[51];通过对转录调节因子SPT15的突变,可以实现其对乙醇和葡萄糖的耐受性[52]。同时,在酵母中构建了含锌指的人工转录因子的随机文库,并鉴定了其中与酵母耐热性、耐药性和渗透性等表型相关的突变子。
3.2 基于σ因子的全局基因表达调控工具
σ因子是DNA依赖型RNA聚合酶全酶的一个亚基,只有RNA聚合酶核心酶与σ因子结合组成全酶时,σ因子才有DNA结合功能[53]。在宿主内存在多种不同的σ因子,每一种σ因子均存在启动子偏好,可以通过引导RNA聚合酶转录不同的基因来实现基因表达水平的调控。例如,在大肠杆菌中,一种特殊的σ因子——σ32可以引导核心酶在热休克基因的启动子处转录,从而使细胞耐受于环境温度的改变。在研究中发现,通过对σ因子的突变,可以改变其启动子偏好性,从而影响胞内一系列基因的转录水平,从而在全局水平上调节转录组。这被视为一种全局性的基因表达调控工具。
目前,在典型原核模式微生物中,基于σ因子的全局基因转录调控工具均已被构建和应用。最早的研究集中于大肠杆菌中。通过对大肠杆菌内必需的持家σ 因子——σ70的编码基因rpoD进行随机突变文库的构建,鉴定并表征得到了3种目标突变子:乙醇耐受性菌株、高产目的化学品菌株(高产番茄红素)以及多种所需的表型结合菌株(同时耐受乙醇和十二烷基硫酸钠)[54]。在枯草芽孢杆菌中,调控应激转录因子σB编码基因sigB的表达可以阻止发酵过程生物膜的有害过度生长并加速细胞在饥饿状态下的扩散;通过敲除σB编码基因sigZ,可以防止芽孢的形成,从而使其更适合于大规模的氨基酸发酵。
3.3 基于表观遗传修饰的全局基因表达调控工具
基于表观遗传修饰的基因表达调控工具通过在宿主基因组中引入DNA 修饰以实现全局基因表达水平的调控。表观遗传机制涉及DNA 甲基化、组蛋白修饰和RNA 沉默等过程,所有这些过程均可以控制基因表达[55-56]。表观遗传机制主要作用于真菌、植物等真核生物,尤其在人类胚胎发育、细胞分化、基因组印记以及癌症等疾病的发展中起作用[57]。在细菌中,研究最多的表观遗传调控机制为DNA甲基化,其通过DNA甲基转移酶对基因组进行修饰[58]。目前,在细菌和真核生物中均发现了腺嘌呤和胞嘧啶甲基化,但已证明胞嘧啶甲基化对真核生物的基因调控影响更大,而腺嘌呤甲基化在细菌中具有更大的调控作用。DNA 甲基化在胞内主要实现两种功能:一种是细胞限制-修饰系统的重要组成部分,用以抵御噬菌体的入侵,胞内限制性核酸内切酶切割来源于入侵噬菌体的或其他外源的未被甲基化的DNA,而宿主本身基因组由于存在甲基化而避免了切割;另一种功能是通过与限制-修饰系统无关的孤儿甲基转移酶发挥全局基因表达调控的功能。目前,在典型模式微生物中,已经有研究表明通过调控基因组修饰相关蛋白(如大肠杆菌DNA 甲基转移酶,Dam)可以调控宿主多种生理过程,如DNA复制、DNA 修复、有氧呼吸、氨基酸代谢、核苷酸代谢、鞭毛生物合成和环境抗逆性[55]。
3.4 基于sRNA的全局基因表达调控工具
此外,一种非编码功能的sRNA——6S RNA也可作为全局基因调控工具[59-60]。6S RNA 是广泛存在于细菌中的转录下调全局调控因子,对于调节环境压力和优化营养匮乏期间细胞的存活至关重要[61]。在细胞生长过程中,6S RNA 逐渐积累,通过与持家σ 因子σ70和应激σ 因子σ38相互作用影响RNA 聚合酶全酶对基因转录的启动子偏好性,调控胞内全局性的基因表达[62]。6S RNA 作为一种全局基因表达调控工具,在大肠杆菌中的表达量可以显著影响细胞的环境抗逆性[59]。
3.5 基于CRISPR的基因表达调控工具
CRISPR(规律间隔成簇短回文重复序列)系统是存在于约40%细菌和90%古细菌的外源DNA抵抗系统,其通常由三部分组成:Cas 蛋白,成熟的CRISPR RNA(crRNA)和与crRNA 部分互补的反式RNA(tracrRNA)[63]。当外源DNA 如噬菌体DNA 侵入细胞时,两种RNA 结合后可以引导Cas 蛋白实现对外源基因的切割。在不同的生物体中发现了多种CRISPR 系统,其中最简单,也是目前应用最广泛的一种是化脓链球菌的Ⅱ型CRISPR系统[64-65]。其Cas蛋白——Cas9可以通过突变沉默其DNA 切割功能,保留其DNA 结合功能,得到dCas9 蛋白。在此基础上,利用CRISPR/dCas9 系统的序列结合特异性可以实现多种基因表达调控功能,将dCas9融合强激活域,可以实现目的基因转录水平的提高,称为CRISPRa;将dCas9 靶向于基因的启动子处,或将dCas9 融合强抑制域,如MXI1, 可以干扰目的基因转录, 称为CRISPRi[66-67]。这些基因表达调控工具可以在不改变目的基因表达框的情况下,实现胞内多基因全局表达水平的改变,提高了代谢工程改造效率。目前,在3种典型模式微生物中,均发展出了广泛应用的基于CRISPR 的基因表达调控工具。随着工具的进一步发展,也已经开发出了基于多种dCas蛋白的基因表达调控工具,如dCpf1,其相较于dCas9 增加了对于RNA 的编辑功能,提高了能够同时调控的基因数量[68]。
4 响应特定信号的基因表达调控工具箱
静态基因表达调控工具可以实现合成生物学对于基因表达的基本要求,并在代谢工程中通过调节基因表达量平衡胞内各代谢途径的通量。但是,当静态基因表达调控工具存在局限时,如目的基因的表达或中间代谢产物对细胞生长造成损害或加剧细胞代谢负荷时,实现精确的基因表达动态调控显得越来越重要。经典的基因表达调控元件中,仅有很少的工具,例如诱导型启动子可以实现对于基因的动态表达调控,但启动子数量的有限、诱导剂的额外添加、较小的动态范围以及基础表达的泄露使其越来越不能满足合成生物学研究与代谢工程应用的需要,亟待发展出新型的可响应多种特定信号的基因表达动态调控工具[2,43]。
在胞内,动态的基因表达调控是天然代谢产物的普遍调控方式。基于此,目前新发展出了一系列新型的天然基因表达调控工具。例如,通过天然代谢产物对基因表达的反馈调节开发了生物传感器(biosensor),通过宿主间基于化学信号交流导致特定基因表达的现象开发了群体响应分子开关。此外通过组合天然发现的分子工具或通过人工改造已有分子工具的生物学特性,开发出了一系列天然不存在的人工基因表达调控工具。这些工具及其组合的使用可以实现类似于天然的基因表达调控功能,在缓解细胞代谢负荷、胞内中间代谢产物自动控制、解偶联细胞生长与生产过程等方面具有广阔的应用前景,成为未来研究和开发的重要方向之一。
4.1 基于光遗传学的基因表达调控工具
经典的动态基因表达调控元件——诱导型启动子通过诱导剂的添加可以实现基因表达的激活,但是当需要停止基因表达时,很难立即移除细胞培养环境中的诱导剂。因此,需要开发一些能够实现输入信号瞬间开启或移除的基因表达调控工具。在所有的输入信号中,光是一种理想的基因表达调控信号,其可以被瞬间开启或关闭[69]。因此,有必要开发基于光遗传学的基因表达调控工具。在自然界中,天然存在多种光调控的基因表达元件,通过光信号的输入,可以实现胞内蛋白的相互作用或基因转录的开启[70-72]。光诱导基因的转录一般基于感光蛋白结构的变化,在有特定波长光照射的情况下,其可以将光信号转化为基因表达信号,当移除光源时,感光器会发生暗还原,恢复原始状态。基于这一原理,最初在神经元的控制领域构建了光遗传学工具,用以在完整的哺乳动物神经组织内实现神经元刺激或抑制的精确时空调控[73-76]。
目前光遗传学工具箱已广泛扩展应用至所有的典型模式微生物,并在离子选择性、光谱灵敏度和时间分辨率等方面进行了优化,以用于合成生物学研究和代谢工程改造。在大肠杆菌中,基于蓝细菌植物色素8(Cph8)构建了红光依赖的基因表达调控工具,基于蓝细菌膜相关组氨酸激酶CcaS 及其调节蛋白CcaR 构建了红光依赖的基因表达调控工具,使工程大肠杆菌能够独立感应红光和绿光,实现多基因表达的精确时空调控[77-78]。这套光遗传调控系统进一步被引入至枯草芽孢杆菌,通过优化途径基因表达,在枯草芽孢杆菌内成功实现了超过70 倍的基因表达激活[79]。在酿酒酵母中,基于一种蓝光激活的光、氧、电压结构域(lightoxygen-voltage sensing domain,LOV)光敏蛋白,EL222 和相应光敏转录因子结合启动子PC120,成功构建了两个酵母光遗传基因表达工具OptoEXP 和OptoINVRT,分别用于蓝光依赖的基因表达激活与基因表达抑制,并成功应用于代谢工程中提高酿酒酵母异丁醇和2-甲基-1-丁醇的产量[73](图2)。
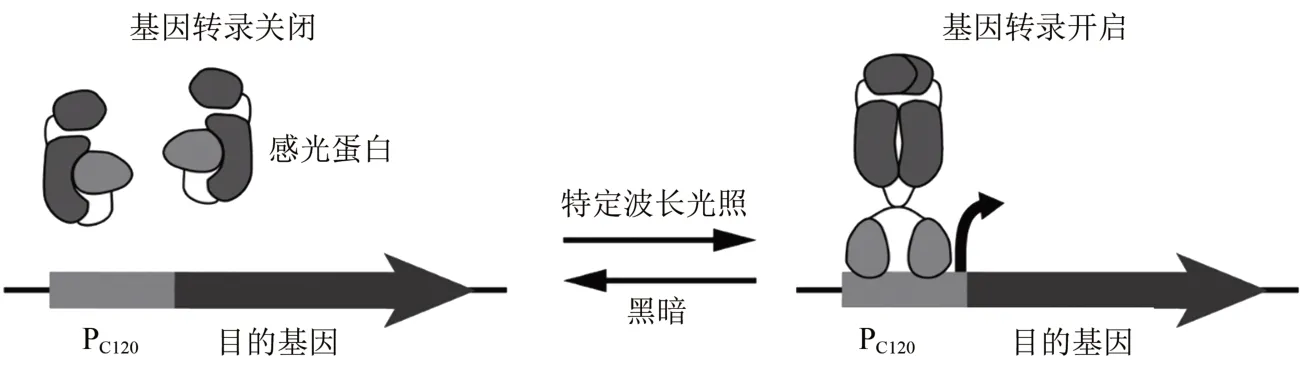
图2 基于光遗传学的基因表达调控工具Fig.2 Optogenetic-based regulation tools for gene expression
4.2 基于蛋白降解标签的基因表达调控工具
在目前的基因表达精确调控工具中,大部分工具都集中在对转录和翻译过程的调控,这些工具适用于大多数情况。但是当构建的遗传回路需要更短的响应时间时,在转录和翻译水平上的调控可能造成遗传回路的失效,而在蛋白水平上对蛋白丰度的调控理论上更加迅速[80]。在自然界中天然存在一些蛋白丰度调控信号,其中蛋白降解标签由于组成和操作的简便性,已经应用于基因表达调控工具的开发[81-83]。目前,相关工具的应用大多在大肠杆菌中进行。例如,在大肠杆菌中通过将蛋白降解的端与CRISPRi 结合,开发了能够应用于必需基因鉴定和抗生素发现的基因表达调控工具[84];将蛋白降解标签用于构建大肠杆菌中具有正交性的遗传回路,实现了基因表达200倍的抑制与8~190 倍的激活[85];在大肠杆菌中构建了基于病毒蛋白酶与蛋白水解信号的代谢开关,进一步开发了包括3种动态调节回路的合成生物学工具箱,实现了代谢通量的快速响应[80]。
4.3 基于群体响应的基因表达调控工具
当工程细胞外源基因的表达不利于细胞的生长或中间代谢产物具有细胞毒性时,将细胞的生长过程与生产过程解偶联是最佳的解决方法之一,即先使细胞生长至合适状态,再开启生产过程[86]。在之前介绍的动态基因表达精确调控工具中,基因表达状态的改变均依赖于人为输入信号,如诱导物的添加、光照条件的改变等。这就需要对细胞状态具有良好的判断,增加了发酵生产的不确定性与复杂性[42]。解决方法是构建能够自动判断细胞所在群体状态和细胞密度的基因表达调控工具以自动开启生产过程[87]。天然存在的理想的细菌交流系统,即细菌群体响应系统,其通过细胞外化学信号的释放与接收,响应细胞密度的变化。例如,当环境中的细胞密度超过某一阈值时,胞内特定基因的表达状态会发生,如特定基因表达的激活或抑制。
基于这一原理,已经在两种典型模式原核生物中建立并优化了基于群体响应的基因表达调控工具。例如,在大肠杆菌中,基于LuxR 群体响应系统的基因表达调控工具可在细胞生长中后期将TCA 循环的通量转向异丙醇生产,基于EsaR 群体响应系统的基因表达调控工具可在细胞生长中后期将糖酵解通量转向葡萄糖二酸生产[88-89]。在枯草芽孢杆菌中,使用基于Phr60-Rap60-Spo0A 的双功能QS 系统构建遗传基因回路可将维生素K2产量提高40 倍[90]。由于真核微生物与原核微生物在胞内环境、转录元件结构等方面存在巨大差异,因此将细菌中的群体响应基因表达调控元件应用到酿酒酵母中仍然具有挑战性。但是,目前也已经设计出了基于酵母细胞间信息素通讯的群体感应系统,并成功应用于提高酿酒酵母对羟基苯甲酸等化学品的产量[91]。
4.4 基于生物传感器的基因表达调控工具
在发酵过程中,对于工程菌株的理想要求是其能够像野生型菌株一样,自动监测环境和胞内代谢物平衡状态,并自动进行不同代谢途径之间代谢流量的调节。但在工程菌株生产高附加值化合物的过程中,目的途径往往被过表达或引入的异源代谢途径会合成原有代谢网络动态调控范围以外的代谢产物。这使得细胞不能对这些途径进行自动、合理的动态调控以减轻细胞代谢负荷。为了最大限度地减少与这种生物合成过程相关的负担,最近的一些研究通过模仿天然基因调节网络,开发出了可以响应特定代谢产物并实现细胞自主调节的基因表达调控工具,称为基于生物传感器的基因表达调控工具[1]。生物传感器可以将特定代谢物浓度信号作为输入并将其转换为对特定基因的转录调控,在典型模式微生物中均开发出了这种基因表达工具,并且其在合成生物学研究和代谢工程领域应用十分广泛。
基于生物传感器的基因表达调控工具首先可以应用于对胞内代谢途径的自动调控。例如,在大肠杆菌中,开发了基于丙二酰-CoA 生物传感器的基因表达调控元件,通过自动调控脂肪酸合成关键基因的表达实现了细胞生长和产物合成间的平衡[92];在枯草芽孢杆菌中,构建了基于GlcN6P生物传感器的基因表达调控元件,可以在检测到细胞内GlcN6P 积累后激活下游基因表达同时抑制上游基因表达[68]。基于生物传感器的基因表达调控工具还可以用于实现细胞合成“成瘾”策略。例如,在大肠杆菌中通过使用甲羟戊酸生物传感器调控细胞必需基因的表达,可以实现对培养体系内产物合成亚群生长的激励与产物非合成亚群生长的抑制[93]。此外,基于生物传感器的基因表达调控工具可以将细胞表型与产物合成偶联,从而获取高产菌株[94-95]。例如,在大肠杆菌和酿酒酵母中,使用酪氨酸和类异戊二烯生物传感器增加菌株突变率并根据荧光蛋白表达强度筛选高产菌株[96];在酿酒酵母中,将芳香族氨基酸生物传感器与细胞抗生素抗性相偶联,通过定向进化得到黏康酸高产菌株[97]。
5 结论与展望
目前,典型模式微生物基因表达精细调控工具已经围绕中心代谢各个层面进行了补充、增强与发展,包括基因水平、转录水平、翻译水平和蛋白水平,实现了工具的多样化和创新性(表1)。同时,新型的基因表达调控工具已经在一定程度上实现了理想基因表达调控元件的特征,如严格控制的泄露率、便于调节的动态模式、宽泛的响应区间、良好的正交性和稳定的信号响应性等。此外,随着系统生物学和计算分析技术的进一步发展,研究人员发现了越来越多性能强大的天然细胞基因表达精细调控工具,并且已经用于动态基因表达精细调控。同时,根据日益丰富的生物学信息开发功能更强大的基因表达精细调控元件也是未来发展的方向之一。性能更强大的基因表达调控工具可以进一步促进合成生物学和代谢工程的发展,如使工程菌株工程菌能够响应监测到代谢压力而自动精确调节基因表达,在实现胞内资源平衡、防止代谢毒性和抑制低产亚群等方面具有广阔的前景。此外值得注意的是,在3种典型模式微生物中,由于酿酒酵母基因表达调控机制,如转录调控元件结构、群体响应机制等相较于原核生物更加复杂,且其基因改造效率远低于大肠杆菌与枯草芽孢杆菌,使得在酿酒酵母中基因表达调控元件的发展相对于典型模式原核微生物来说仍然相对缓慢。随着系统生物学和生物信息学的快速发展和对真核生物基因表达调控机制的了解更加深入,可以实现酿酒酵母基因表达调控元件的进一步拓展与完善。

表1 模式微生物新型基因表达调控工具Tab.1 Regulatory tools of gene expression for model microorganisms
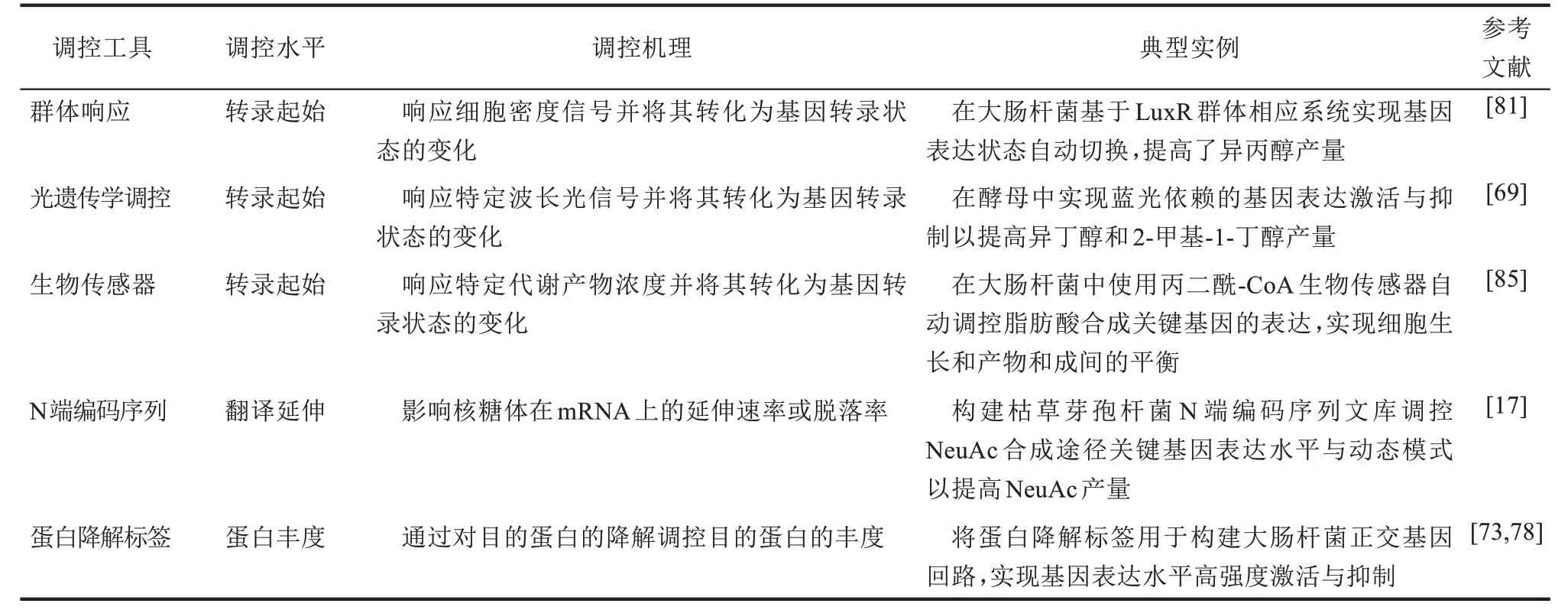
续表
同时,目前的基因表达调控工具仍然存在一些亟待解决的问题。首先是响应元件的特异性,如在自然界中得到的天然生物传感器,其往往可以同时响应一类化合物且极难改造和优化,这使得复杂的胞内环境对其存在极大的干扰。其次,当前的基因表达调控工具的开发存在不同的标准,如何通过大数据的分析或构建统一的参考标准将不同基因表达调控进行强度或动态模式的标准化是一项巨大的挑战。最后,当前的新型基因表达调控工具大多停留在实验室层面,特别是响应多种信号的动态调控元件,如何实现其从概念验证到工业化大规模应用的成功过渡是未来发展所注重的问题。通过机器学习算法对目前已有数据的整合分析,为基因表达调控工具的应用提供预测与参考是有望解决上述问题的重要途径。
猜你喜欢
理财周刊(2022年4期)2022-04-30
中国特种设备安全(2022年1期)2022-04-26
昆明医科大学学报(2022年1期)2022-02-28
中国畜禽种业(2021年11期)2021-12-09
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
中国医学影像技术(2019年6期)2019-06-24
中国兽医杂志(2019年10期)2019-05-20
中国核电(2017年2期)2017-08-11
安徽医科大学学报(2015年9期)2015-12-16
