
裸盖鱼精子超微结构
2020-10-27 08:33孙静娴陈仕友王茂林李雪杰
广东海洋大学学报 2020年6期
孙静娴,陈仕友,王茂林,王 伟,李雪杰
裸盖鱼精子超微结构
孙静娴,陈仕友,王茂林,王 伟,李雪杰
(大连海洋大学水产与生命学院,辽宁 大连 116023)
【】研究裸盖鱼()精子的结构特征。利用扫描电镜和透射电镜观察裸盖鱼精子的超微结构。【】裸盖鱼精子由头部、中段和尾部组成,无顶体;精子头部与中段无明显区分,呈椭球形,长径1.28~2.00 μm;尾部细长,长20.4~26.3 μm。精子头部主要由细胞核构成,细胞膜有小褶皱并紧密包裹细胞核。细胞核椭球形(长度略小于宽度),结构致密,核内染色质呈高电子密度斑块状且分布较均匀,有核泡。细胞核后部有一约占核直径2/5的植入窝(核凹窝)。中段由中心粒复合体、袖套和线粒体等构成,近端中心粒位于植入窝内,退化为一些微管和小囊泡,远端中心粒(基体)位于植入窝外侧,基体头端由电子致密物质构成,并向后发出轴丝,成为尾部鞭毛的中轴结构;袖套与细胞核后端相连,分布有4个球形线粒体及数量较多的囊泡等,囊泡含有电子密度低的颗粒状物质。尾部细长,鞭毛起始于基体,从袖套腔中伸出;轴丝为鞭毛的中心结构,具典型的“9+2”型双联微管结构,外侧有波浪形侧鳍,侧鳍结构与提高精子受精率有关。
裸盖鱼;精子;超微结构
裸盖鱼()隶属于辐鳍鱼纲(Actinopterygii)鲉形目(Scorpaeniformes)黑鲉亚目(Anoplopomatoidei)黑鲉科(Anoplopomatidae)裸盖鱼属(),为太平洋冷水性底栖鱼类,主要分布于北太平洋沿岸,包括墨西哥北部至白令海峡沿岸、勘察加半岛至日本东北部沿海[1],是北太平洋商业捕捞的重要种类,有较高的商业价值[2]。目前,关于裸盖鱼研究主要集中在繁殖生物学、渔业资源、工厂化养殖、染色体核型、分子生物学等方面。Head等[3]对裸盖鱼生活史参数的研究发现,在研究的天然海域中,50%成熟时,鱼体长(50)和年龄(50)估计值分别为54.64 cm和6.86龄;温度较低水域更易获得较年轻年龄的较大尺寸(50%成熟时)的裸盖鱼。我国于2013年引进裸盖鱼,2015年成功实现人工育苗[4],但关于其雄性生殖发育和精子生理机能方面的相关研究较少,仅见Immermman等[2]研究的最佳精子激活条件及优化的裸盖鱼精子冷冻保存体系,仍需对其受精生物学进行进一步研究。本研究通过观察裸盖鱼精子超微结构,探讨裸盖鱼精子结构特征,为该物种繁殖特性评估及受精生物学研究提供基础资料,为其繁育推广和人工育种过程中的种质资源保护提供参考。
1 材料与方法
1.1 试验材料
采精用裸盖鱼由大连海洋岛海洋集团有限公司提供,为繁殖季节(4月)雄性性成熟亲鱼,体长40.5 ~ 80.6 cm,体质量3.3 ~ 7.9 kg。暂养用水为过滤自然海水,照度低于100 lx,水温维持在6 ~ 8 ℃(制冷冷却),盐度34,pH 8.1,连续充气,溶氧保持在5 mg/L以上。每天投喂冰鲜杂鱼1次,水池清底1次。
1.2 试验方法
1.2.1 精液采集 取精液过程全程避光并避免水分。采精时腹部朝上,轻压腹部,挤出膀胱中的尿液,用医用棉纱擦干鱼体生殖孔附近,待挤出的精液呈较纯的乳白色后,用注射器吸取精液(避免尿液、粪便及血液污染)。将干净精液置于1.5 mL离心管中,用黑色塑料袋包裹,及时放入有冰袋的保温箱,4 h内低温、避光送至实验室。取1滴精液于载玻片,加等量过滤海水稀释,激活精子,分析精子活力。选取活力高、未被污染的精液0.5 mL,加入2 mL预冷的体积分数2.5%戊二醛溶液固定。因鲉亚目三种鱼圆鳞鲉、斑马纹多臂蓑鲉(鲉科)[(Scorpaenidae)]、毒鲉(毒鲉科)[(Synanceiidae)]的分类地位和生活环境与裸盖鱼比较接近,本研究固定液磷酸盐缓冲液(PBS)仿照该文献的PBS配方[5],即0.1 mol/L、pH 7.4的PBS,加25 g/L NaCl、15 g/L蔗糖,调渗透压至约425 mmol/L。固定15 min后,全量更换固定液,于4 ℃下固定保存8 h。
1.2.2 扫描电镜样品制备 固定的样品按文献[6]方法制备。用1.2.1固定液漂洗。分别用体积分数50%、70%、80%、90%叔丁醇梯度脱水,然后用纯叔丁醇重复脱水3次。将样品置于纯叔丁醇中冷冻干燥,然后放入离子溅射(JFC-1100,JEOL公司)中喷金。扫描电镜(JSM-6360LV,日本电子)观察拍照(加速电压15 kV)。
1.2.3 超薄切片样品制备 固定的样品按文献[6]方法制备。样品用20 g/L琼脂糖预包埋,用0.1 mol/L、pH 7.2 ~ 7.4的PBS冲洗。用体积分数1%锇酸固定2 h(20 ℃室温)。经PBS缓冲液冲洗、乙醇梯度脱水、Epon 812 环氧树脂包埋,于37、45、60 ℃恒温箱固化。用超薄切片机(Ultracut E,奥地利)切片,切片经0.04 mol/L醋酸双氧铀和柠檬酸铅双染色法染色。透射电镜(JEM-2000EX,JEOL;加速电压120 kV)观察,AMT Camera System拍照。
2 结果
2.1 精子外形
图1可见,裸盖鱼精子分为头部、中段和尾部,无顶体(图1d、e)。头部长径为1.28~2.00 μm,椭球形,表面可见一凹陷;尾部细长,长20.4~26.3 μm(图1a、b)。
2.2 精子头部超微结构
头部主要由细胞核构成,椭球形(长度略小于宽度)。细胞膜有小褶皱,细胞质少。细胞膜与核膜紧密连接,无其他结构(图1e)。细胞核结构致密,核内浓缩的染色质呈高电子密度斑块状,染色质间隙相互连接呈网状。染色质中存在未完全浓缩、相对疏松的区域,为核泡(图1c)。细胞核上有一处核膜向内凹陷的凹窝,凹窝开口于细胞核底部(细胞核朝向精子中段处),称为植入窝(核凹窝)。植入窝约占核直径的2/5,走向与细胞核长轴平行(图1f、g)。精子头部似有背腹面之分,细胞核无植入窝区域位于腹面,植入窝略偏向背面(图1d),在精子背侧表面形成一个凹陷(图1b)。
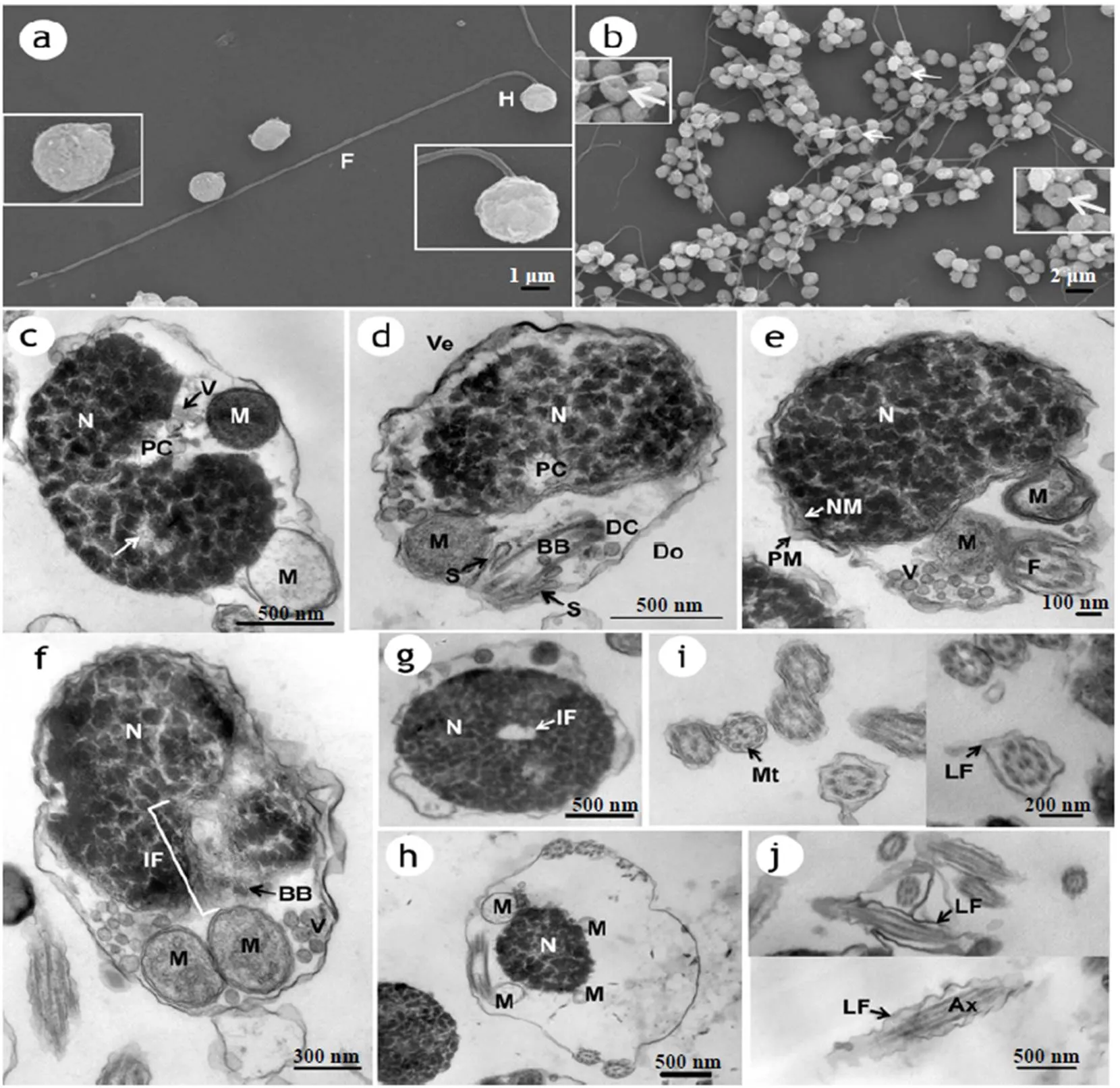
a-b. 扫描电镜下精子外部形态,箭头示凹陷;c-j. 透射电镜下精子内部超微结构;c. 精子头部及中段纵切,近端中心粒退化为微管和小囊泡,箭头示核泡;d. 精子头部及中段纵切,示中心粒复合体、袖套、线粒体结构和精子背腹侧;e. 精子头部及中段纵切,示袖套中的囊泡、鞭毛横切、细胞质膜与核膜结构;f. 精子头部及中段纵切,示细胞核植入窝、中心粒复合体、袖套中的线粒体与囊泡结构;g. 精子头部经细胞核横切,示细胞核植入窝;h. 精子中段横切,示线粒体和细胞核;i-j. 鞭毛横切和纵切,示鞭毛中轴丝、微管和侧鳍结构。Ax,轴丝;BB,基体;DC,远端中心粒;F,鞭毛;H,精子头部;LF,侧鳍;M,线粒体;Mt,微管;N,细胞核;NM,核膜;PM,细胞质膜;PC,近端中心粒;S,袖套腔;V,囊泡;IF.植入窝;Do,背侧;Ve,腹侧
2.3 精子中段超微结构
中段与头部紧紧相连,主要由中心粒复合体、袖套和线粒体等构成(图1d)。中心粒复合体位于头部背侧,包括近端中心粒和基体(远端中心粒)两部分。近端中心粒位于植入窝内,结构不清晰,发生了退化,无典型的中心粒结构,仅有一些微管和小囊泡(图1c、d)。远端中心粒(基体)位于植入窝外侧,头端由电子致密物质构成,并向后发出轴丝,构成尾部鞭毛的中轴结构。鞭毛起始部位(基体)周围有扩大的空腔。袖套位于细胞核后端,由细胞膜向下延伸形成(图1d)。袖套内有约4个球形线粒体(图1h)、数量较多的囊泡及少量的细胞质。线粒体有清晰的双层膜结构和较明显的嵴(图1d-f),囊泡中有电子密度低的颗粒状物质(图1e、f)。袖套短圆筒状,包绕着鞭毛(图1e)。
2.4 精子尾部超微结构
尾部主要由鞭毛构成。鞭毛中心结构是轴丝。基体(远端中心粒)发出的轴丝不断向外延伸,从袖套中伸出,成为细长的鞭毛。轴丝有典型的“9+2”型双联微管结构(图1i)。鞭毛轴丝的外侧有由细胞质膜向两侧扩展成的波浪形侧鳍(图1i、j)。
3 讨论
尹洪滨等[7]认为分类地位越近的鱼类精子结构越相近。Pavlov等[5]研究表明,鲉亚目中三种鱼类圆鳞鲉属圆鳞鲉、鲉科斑马纹多臂蓑鲉和毒鲉科毒鲉的精子均有圆形头部(长度略小于宽度)、中等发达的中段和鞭毛。圆鳞鲉精子中围绕着鞭毛的起始部分有扩大的胞质通道,有大量的线粒体节,鞭毛最长,与其他两种鱼类不同。毒鲉与斑马纹多臂蓑鲉无显著差异。然而,根据头部和中段轮廓(椭圆傅立叶分析),毒鲉又与圆鳞鲉和斑马纹多臂蓑鲉有明显的分离趋势,认为鲉亚目中三种鱼类精子形态的比较数据与相应类群的系统发生模式有关,同时,精子形态描述的资料还可用于评估该物种的繁殖特性。Jamieson[8]也认为精子各结构的形态特征可提供鱼类进化与系统学的证据。因此,研究裸盖鱼精子超微结构,除可为裸盖鱼生殖进化研究提供基础资料,还可为提高裸盖鱼人工授精、育种的成功率,保护种质资源等实际问题提供参考。
3.1 精子头部
裸盖鱼精子头部呈椭球型,在扫描电镜下未进行详细测量,无法确定具体长宽比例。在透射电镜下精子内部超微结构显示,构成精子头部的细胞核也呈椭球形(长度略小于宽度),与Pavlov等[5]描述的鲉亚目中三种鱼类精子头部形状类似。
大多数硬骨鱼类精子细胞发育需经历早期、中期和后期3个主要阶段。发育过程中细胞核中的大部分染色质高度浓缩,少部分非浓缩的染色质形成核泡,核泡的有无及其结构有种的特异性[9]。舒琥等[10]认为核泡只是在致密染色质中相对疏松的区域。刘利平等[11]认为,在受精过程中,精子的不断运动可能对细胞核内遗传物质造成冲击,核泡可能有缓冲机械压力的作用。裸盖鱼精子头部细胞核染色质中存在少量核泡,其生理功能需进一步研究。
硬骨鱼类精子头部细胞核的后端一般有植入窝(核凹窝),精子头部形态和植入窝结构特点有硬骨鱼属种差异,并与受精方式相关。同属鱼类头部形态因受精方式不同而不同,体外受精类型鱼类精子头部为典型圆形,而体内受精类型则为稍长形;体内受精类型鱼类头部植入窝的发达程度超过体外受精类型[12-13]。裸盖鱼为体外受精型,精子头部保持最原始的圆形,细胞核植入窝约占核直径的2/5,中等发达。头部有背腹之分,细胞核无植入窝区域位于腹侧,植入窝区域附近的中心粒复合体位于头部的背侧,与大黄鱼()精子头部结构类似[14]。
3.2 精子中段
精子中段有无近、远端中心粒,中心粒排列方式可作为种之间相互区别的依据[9]。裸盖鱼近端中心粒位于精子细胞核的植入窝内,但结构不清晰,发生了退化,不具有典型的中心粒结构,仅剩一些微管和小囊泡,与尼罗罗非鱼鱼 ()[15]类似;褐菖鲉()在精细胞变态为精子的过程中发生近端中心粒消失现象,被看作是精子成熟的最后标志[16]。而舒琥等[10]认为精子近端中心粒排列方式的差异可能由切片角度不同所造成,精子近端中心粒的排列方式是否可作为硬骨鱼类种分类特征还有待进一步研究。因此,裸盖鱼近端中心粒的退化机制需进一步研究。
裸盖鱼精子的远端中心粒(基体)位于核植入窝外侧,类似于黄姑鱼()[17]、大黄鱼[14]和短吻鮠(Wu)[18]。Pavlov等[5]发现圆鳞鲉精子中围绕鞭毛的起始部分有扩大的胞质通道。裸盖鱼精子围绕鞭毛起始部位(基体)周围也有扩大的空腔,是否存在扩大的胞质通道还需进一步研究。
裸盖鱼精子袖套中有4个线粒体,单层排列。除线粒体外,还分布较多囊泡,囊泡中含有电子密度低的颗粒状物质。舒琥等[10]认为尚不能确定囊泡结构的作用及其所在位置是否可作为硬骨鱼类种间的鉴定依据。尤永隆等[15]认为在精子细胞的成熟过程(精子形成)中,在染色质的浓缩过程中,核的体积和表面积均大为缩小。可能是因为核膜一部分构成了囊泡的膜,再将细胞核内某些电子密度低、未参与染色质浓缩的物质包在其中排出细胞核,最终排出精子细胞。裸盖鱼囊泡来源的具体机制和作用还需进一步研究证实。
3.3 精子尾部
裸盖鱼尾部细长,鞭毛轴丝外部具有由细胞质膜向两侧扩展而成的侧鳍。硬骨鱼类精子均有细长的尾部,这与其多于水中受精的方式相适应。其鞭毛结构有种属差异性,鞭毛长度,侧鳍有无及发达程度可作为硬骨鱼类分类依据。
鞭毛是精子的运动器官,侧鳍的有无影响精子的游泳速率,从而影响受精效率,不同学者有不同的看法:Stoss[19]认为侧鳍可能改善精子鞭毛的游泳速率,有利于提高受精率;尹洪滨等[7]认为发达的侧鳍为精子的快速游动提供保障,并在其灵活准确的定向游动中发挥重要作用;Afzelius[20]、张汉峰等[18]则认为侧鳍与精子游泳速率的提高无多大关系。笔者认同尹洪滨等[7]的观点,认为体外受精鱼类的精子进入水体后,需保持平衡,维持快速有序游动,避免互相缠绕,以利于提高受精率。因此,侧鳍与精子游泳速率的有一定关系,同时可为精子提供一定的浮力,并保持精子游动的平衡,通过快速、分散、积极游动(鞭毛有一定的长度,适当数量的线粒体供能),提高受精率。
[1] BEAMISH R J, CHILTON D E. Preliminary evaluation of amethod to determine the age of sablefish ()[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1982, 39(2): 277-287.
[2] IMMERMMAN D A, GOETZ F W. The activation and cryopreservation of sablefish () sperm[J]. Aquaculture, 2014, 430: 211-217.
[3] HEAD M A, KELLER A A, BREADBURN M. Maturity and growth of sablefish,, along the U.S. West Coast[J]. Fisheries Research, 2014, 159: 56-67.
[4] 曹栋正, 陈四清, 王贞杰, 等. 裸盖鱼幼鱼工厂化养殖试验[J]. 渔业现代化, 2016, 43(3): 23-28.
[5] PAVLOV D A, EMEL’YANOVA N G. Comparative analysis of spermatozoa morphology in three fish species from the suborder Scorpaenoidei[J]. Journal of Ichthyology, 2018, 58(2): 226-238.
[6] 张东升, 周玮, 孙静娴, 等. 光镜与电镜技术及其在水生生物上的应用[M]. 北京: 中国农业科学技术出版社, 2018: 8.
[7] 尹洪滨, 孙中武, 于波. 乌苏里拟鲿()精子的超微结构[J]. 广东海洋大学学报, 2009, 29(6): 28-31.
[8] JAMIESON B G M. Fish evolution and systematics: evidence from spermatozoa[M]. London: Cambridge University Press, 1991.
[9] 杨万喜, 应雪萍, 竺俊全, 等. 硬骨鱼类精子发生及其在系统演化研究中的应用前景[J]. 东海海洋, 2000, 18(3): 53-58.
[10] 舒琥, 谭嘉敏, 赵会宏, 等. 黄鳍东方鲀精子的超微结构[J].广东海洋大学学报, 2008, 28(6): 9-13.
[11] 刘利平, 王武, 赵雷蕾, 等. 江黄颡鱼精子的超微结构[J]. 上海水产大学学报, 2004, 13 (3): 198-202.
[12] 刘雪珠, 杨万喜. 硬骨鱼类精子超微结构及其研究前景 [J]. 东海海洋, 2002, 20 (3): 32-37.
[13] 郑学斌, 张清科, 乐韵, 等. 香鱼() 精子的超微结构及其与鲤形目及鲑形目其他鱼类精子结构的比较研究[J]. 海洋与湖沼, 2018, 49(4): 866-872.
[14] 尤永隆, 林丹军. 大黄鱼精子的超微结构[J]. 动物学报, 1997, 43(2): 119-126.
[15] 尤永隆, 林丹军. 尼罗罗非鱼精子形成中核内囊泡的释放[J]. 动物学报, 1998, 44(3): 257-263.
[16] 林丹军, 尤永隆. 褐菖鲉精细胞晚期的变化及精子结构的研究[J]. 动物学研究, 1998, 19(15): 359-366.
[17] 胡谋, 苗亮, 李明云, 等. 黄姑鱼()与大黄鱼()精子超微结构的观察与比较[J]. 生物学杂志, 2014, 31(2): 1-4.
[18] 张汉峰, 魏刚, 黄林, 等. 短尾鮠精子细胞和精子的超微结构研究[J]. 西南农业大学学报 (自然科学版), 2005, 27(5): 696-699.
[19] STOSS J. Fish gamete preservation and spermatozoon physiology[M] // HOAR W S, RANDALL D J, RANDALL E M. Fish physiology (8B). New York: Academic Press, 1983: 307-308.
[20] AFZELIUS B A. Fine structure of the garfish spermatozoon[J]. J Ultr Res, 1978, 64: 309-324.
Ultrastructure of Spermatozoon of
SUN Jing-xian, CHEN Shi-you, WANG Mao-lin, WANG Wei, LI Xue-jie
(,,116023,)
To study the spermatozoon structure of sablefish,.The ultrastructures of the of sablefish () spermatozoon were observed using scanning electron microscopy (SEM) and transmission electron microscopy (TEM).The spermatozoon consist of a spheroidic head with diameter 1.28 ̶2 μm, a moderately developed midpiece and flagellum (long and thin, 20.4 ̶26.3 μm in length). There is no acrosome on the head, and the head is not distinct from the midpiece. The nucleus is the main structure of the head. The cell membrane closely surrounds the nucleus, which has little folds. The nucleus was spheroid in shape, its length is slightly smaller than its width. The chromatin in the nucleus is highly condensed into electron-dense plaques and is separated by electronlucent zone. Several nuclear vacuoles occur. The nucleus near the flagellum segment is a depression,and it forms a moderately deep nuclear fossa. The midpiece of the spermatozoon contains the centriolar complex, the sleeve, and mitochondria. The proximal centriole is located inside the nuclear fossa, degenerating into microtubules and vesicles. The distal centrioles locating outside the nuclear fossa are differentiated into a basal body and formed into an axonem. The sleeve is linked with the end of the nucleus. There are four round mitochondria and numerous vesicles consist of low electron-dense particles arranged in the sleeve. The flagellum of the tail derived from basal body stretched out of the central space of the sleeve. The axoneme of the flagellum has the typical “9 + 2” structure. The wavy lateral fins are distributed on both sides of the flagella. The lateral fin is probably used to improve the fertilization ability.
; spermatozoon; ultrastructure
Q248;Q954.43
A
1673-9159(2020)06-0130-05
10.3969/j.issn.1673-9159.2020.06.017
孙静娴,陈仕友,王茂林,等. 裸盖鱼精子超微结构[J]. 广东海洋大学学报,2020,40(6):130-134.
2019-10-23
辽宁省重点研发计划(2018203004);辽宁省海洋研究院项目(2018-CY-33)
孙静娴(1978-),女,实验师,研究方向为海产经济动物发育生物学。E-mail:sjingx@163.com
王茂林(1980-),男,副教授,研究方向为鱼类生理生态学。E-mail:wangmaolin@dlou.edu.cn
(责任编辑:刘庆颖)
猜你喜欢
实用检验医师杂志(2019年3期)2019-11-01
人人健康(2019年10期)2019-10-14
特种经济动植物(2019年9期)2019-01-08
数码世界(2018年1期)2018-12-23
科学家(2017年20期)2017-11-10
现代检验医学杂志(2016年3期)2016-11-15
天然产物研究与开发(2016年6期)2016-06-05
中国病理生理杂志(2015年8期)2015-12-21
中国学术期刊文摘(2015年16期)2015-11-01
中国医疗美容(2015年4期)2015-04-27
