
广东大鹏湾北部海域春秋季鱼类群落结构及其与环境因子关系
2020-10-27 08:59邱俊文陈春亮曾嘉维王学锋
广东海洋大学学报 2020年6期
张 静,邱俊文,陈春亮,曾嘉维,王学锋
广东大鹏湾北部海域春秋季鱼类群落结构及其与环境因子关系
张 静1,2,邱俊文1,陈春亮3,曾嘉维1,2,王学锋1,2
(1. 广东海洋大学水产学院,广东 湛江 524088;2. 南方海洋科学与工程广东省实验室(湛江),广东 湛江 524025;3. 广东海洋大学海洋与环境监测中心,广东 湛江 524088)
【】研究大鹏湾北部海域鱼类群落结构及其与环境因子的关系,为掌握该海域的资源现状及其优势种群的变动规律、促进资源增殖提供基础。利用2018年秋季(10月)、2019年春季(3月)的资源生态调查数据,计算鱼类群落指数,并运用典范对应分析(CCA)研究鱼类群落结构与环境因子之间的关系。该海域共捕获鱼类72种,隶属于10目35科59属。其中日本海鰶()、二长棘鲷()、褐蓝子鱼()和颈斑鰏()为两个季度共同优势种。秋季鱼类丰富度指数()、均匀度指数()及多样性指数(′)均高于春季。秋季平均分类差异性指数(Δ+)均值为35.97,其中调查站位S2、S4、S10和S11低于95%置信漏斗区间的下边界,分类差异变异指数(∧+)均值为109.85,表明群落受到一定的外界干扰;春季Δ+均值37.69,∧+均值为100.19,所有站位均处于95%置信漏斗区间内,群落基本稳定。CCA分析表明,盐度、水深是秋季显著影响鱼类分布的重要环境因子,2分别为0.75、0.70,温度为春季显著影响鱼类分布的重要环境因子(2= 0.72)。秋季鱼类群落结构受环境因子变化影响较大,第一、二轴分别解释了13.32%、12.44%的物种分布情况,春季则为16.91%、10.93%。广东大鹏湾北部海域鱼类现存物种资源量秋高春低,其群落结构在秋季受一定的外界干扰,春季基本稳定。
大鹏湾北部海域;鱼类;多样性;群落结构;典范对应分析(CCA)
大鹏湾北部海域位于广东省深圳市大鹏湾西北部,深圳市盐田区境内,西起沙头角保税区、东至正角咀,处于深圳和香港之间,是天然的半封闭港湾。由于存在陆地和香港诸岛作为天然屏障,该港区风浪较小,浪高低于0.7 m,潮汐属不正规半日混合潮。该海域不仅包含了全国集装箱吞吐量最大的单一港区——盐田港,也是目前中国大陆远洋集装箱班轮密度最高的单个集装箱码头,同时也是重要的渔业水域,在港口航运、海洋游钓观光、渔业资源保护等方面均发挥着重要的作用。随着近几年经济的高速发展,港区建设和港口运行必然会给该海域的海洋生物群落结构带来一定程度的影响。
鱼类作为海洋生态环境中最重要的一部分,在物质循环、能量流动中发挥着重要作用。研究其群落结构和功能可以反映该海域的生态系统质量状况,是评估海域生态质量的一项重要指标[1]。目前关于大鹏湾北部港口海域渔业资源的相关研究未见报道,仅在大鹏半岛[2]和鹅公湾[3]两个邻近海域有相关研究。鉴于此,本研究利用2018年10月(秋)、2019年3月(春)两个季度在大鹏湾北部海域进行的渔业资源调查数据,对该海域的鱼类群落结构特征及其与环境因子的关系进行分析,为今后评估该海域的健康状况及鱼类资源的开发利用和保护等提供基础。
1 材料与方法
1.1 站点布设与样品采集
2018年10月17日(秋季)和2019年3月18日(春季)于广东深圳大鹏湾北部海域完成为期1天的鱼类生物资源调查,共设置12个站位(图1)。调查船为“粤汕尾渔11092”虾拖网渔船,船长15.59 m,宽5.6 m,型深2.9 m,主机功率135 kW。所用网具为虾拖网,每张上纲长2.7 m,网口网目大3.0 cm,网囊最小网尺寸2.0 cm。平均捕捞作业时间0.5 h,平均水平拖网航速3 kn。准确记录每一站点起、放网位置和时间,同步采集水深、水温、溶解氧、pH和盐度等5个环境因子,其中水深采用SM-5A型测深仪测定,其余环境因子均采用便携式水质分析仪(YSI Professional Plus)测定。依据《海洋调查规范》[4]GB/T12763.6—2007的操作要求采集和分析渔获物样品。
1.2 鱼类鉴定
参照《南海鱼类检索·上册》[5]和NELSON分类系统对鱼类物种组成进行分类鉴定后,分别进行计数及体长、体质量的测量,体长测量精度精确到1 mm,体质量精确到0.1 g。

图1 广东大鹏湾北部海域调查站位
1.3 数据处理
1.3.1 多样性指数 传统的多样性指数可较好反映物种丰度、均匀度,故选取物种数、Margalef种类丰富度指数、Shannon-Wiener多样性指数、Pielou种类均匀度指数等指数对大鹏湾北部海域的鱼类群落结构特征进行分析。
此外,为更全面地分析、量化群落的分类多样性及分类差异性状况,选取分类多样性指数中的平均分类差异指数(Average taxonomic distinctness, Average TD)Δ+和分类差异变异指数(Variation in taxonomic distinctness, Variation TD)∧+表征大鹏湾北部海域鱼类生物种类组成分类阶元的多样性。各指数计算公式为:
Margalef种类丰富度指数[6],

Shannon-Wiener多样性指数[7],

Pielou种类均匀度指数[8],

平均分类差异指数Δ+[9],
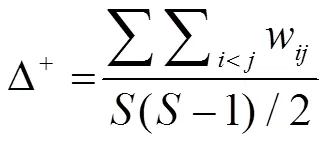
分类差异变异指数∧+[10],
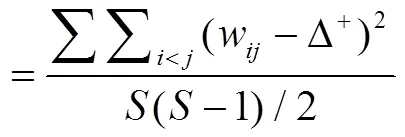
式中,为种类数,为总尾数,P为第种鱼的数量占总渔获数量所比例,w为第和个种类在分类系统树状图中的分支路径长度,采用默认值(即最长的分类路径为100,其余不同分类水平之间的分类路径大小相同)进行计算。以上各指数均采用多元统计软件PRIMER 7软件包完成。
1.3.2 物种优势度 采用相对重要性指数[11](Index of relative importance,IRI)评价鱼类在群落中的重要性,计算公式如下:
IRI =(+)××104, (6)
式中,为某一种类的尾数占总尾数的百分比,为某一种类的质量占总质量的百分比,为某一种类出现的站点数占调查总站点数的百分比。根据程济生等[12]对IRI指数的划分,当IRI > 500时,该种为优势种,100~500为常见种,10~100为一般种,1~10为少见种,小于1为稀有种。
1.3.3 鱼类群落与环境因子的关系 对各站位的鱼类资源密度使用降趋势对应分析(detrended correspondence analysis,DCA)方法判别选用线性模型(Redundancy Analysis,RDA)还是单峰模型(Canonical Correspondence Analysis,CCA)[13]分析物种分布与环境因子关系,根据DCA排序中“梯度之长度量lengths of gradient”项的前四个轴的值进行判断,如果前4个轴的数值最大值超过4,选用单峰模型(CCA);最大值不超过3,则选用线性模型(RDA);介于3 ~ 4之间,两种模型均可以采用。通过蒙特卡洛置换检验(Monte Carlo permutation test)分析物种分布与环境因子相关的显著性,置换循环次数为999[14]。
2 结果与分析
2.1 种类组成
秋、春两季,大鹏湾北部海域鱼类资源调查中共鉴定鱼类72种(表1),隶属于10目35科59属。其中秋季捕获鉴定鱼类54种,隶属于8目26科44属(表2),种类数量占秋、春两季总种类数的75%;春季捕获鉴定鱼类39种,较秋季少,隶属于10目24科33属(表3),种类数量占秋、春两季总种类数的54.17%。

表1 大鹏湾北部海域鱼类资源
从分类上看,两个季度调查均以鲈形目(Perciformes)种类最多,分别占当季度鱼类种类数的64.81%、66.67%。秋季调查鱼类仅有8目54种,而春季较秋季增加了海龙目(Syngnathiformes)和鲇形目(Siluriformes)2目共2种。春季鱼类种类数量下降,较秋季减少了15个物种。

表2 大鹏湾北部海域秋、春季鱼类群落的种类构成
2.2 多样性指数特征
2.2.1 传统物种多样性指数特征 秋、春季大鹏湾北部海域鱼类群落的物种多样性指数如表3所示。秋季,在各站位的变化范围为1.82 ~ 2.30,平均值为2.02;在各站位的变化范围为2.18 ~ 4.37,平均值为3.23;在各站位的变化范围为0.53 ~ 0.89,平均值为0.77。春季,在各站位的变化范围为0.99 ~ 2.03,平均值额为1.52;在各站位的变化范围为1.74~3.11,平均值为2.37;在各站位的变化范围为0.42 ~ 0.87,平均值为0.72。各传统物种多样性指数季节平均值均为秋季大于春季。

表3 大鹏湾北部海域秋、春季鱼类群落多样性特征
2.2.2 分类学多样性指数特征 根据珠江口物种名录,分别计算得到秋、春季大鹏湾北部海域鱼类群落的平均分类差异指数(Δ+)和分类差异变异指数(∧+)的理论平均值如表3,95%置信漏斗曲线如图2、图3所示。可以看出,秋季,鱼类平均分类差异指数(Δ+)介于30.90~38.82之间,平均值为35.97,其中S2、S4、S10和S11这4个站位低于95%置信漏斗区间的下边界;分类差异变异指数(∧+)介于84.38 ~ 125.39之间,平均值为109.85,全部站位均处于95%置信漏斗区间内。春季,鱼类平均分类差异指数(Δ+)介于34.46 ~ 40.82之间,平均值为37.69,全部站位均处于95%置信漏斗区间内;分类差异变异指数(∧+)介于83.30 ~ 117.06之间,平均值为100.19,全部站位均处于95%置信漏斗区间内。

图2 2018年秋季Δ+和∧+的95%置信区间漏斗图

图3 2019年春季Δ+和∧+的95%置信区间漏斗图
2.3 优势度及优势种
两个季度调查的鱼类优势种及资源密度、丰度情况如表4所示。秋季有7种优势种,其中颈斑鰏()在所有站位均有出现,其资源密度和丰度分别为212 kg/km2和1 438尾/km2,其IRI指数最高,为3 851;春季有4种优势种,其中二长棘鲷()在所有站位均有出现,其资源密度和丰度分别为233 kg/km2和5 431尾/km2,其IRI指数为2 957,而日本海鰶()的IRI指数最高,为3 356。两个季度的共同优势种有4种,分别为日本海鰶()、二长棘鲷()、褐蓝子鱼()和颈斑鰏()。
2.4 鱼类群落与环境因子的关系
对秋、春两个季度DCA进行排序,各季度DCA排序前四个轴的最大值均小于3,因此采用基于线性模型的冗余分析(RDA)方法分析鱼类群落与环境因子(水温、盐度、、DO、pH)的关系(图4),物种分布与环境因子的相关显著性检验见表5。
秋季,盐度(31.91 ~ 32.62)对鱼类群落结构影响具有极显著的统计意义(< 0.01),水深(11.0 ~ 18.5m)对鱼类群落结构影响具有显著的统计意义(< 0.05);东侧海域鱼类群落结构与盐度呈正相关,而西侧海域鱼类群落结构与水深呈正相关;盐度与水深之间无相关关系(> 0.05)。5个环境因子对鱼类空间格局的形成贡献率为45.93%,第一排序轴共解释了13.32%的物种分布情况,第二排序轴共解释了12.44%的物种分布情况。
春季,水温(19.8 ~ 20.3 ℃)对鱼类群落结构影响具有极显著的统计意义(< 0.01),其余环境因子的影响无统计意义(> 0.05);东侧海域鱼类群落结构与水温呈负相关,而中、西侧海域鱼类群落结构与水温呈正相关。5个环境因子对鱼类的空间格局的形成贡献率为51.18%,第一排序轴共解释了16.91%的物种分布情况,第二排序轴共解释了10.93%的物种分布情况。

表4 秋、春季优势种资源密度、丰度及IRI指数

虚线表示对鱼类群落的影响具统计意义的环境因子(P < 0.05),实线表示影响不具统计意义(P > 0.05).

表5 大鹏湾北部海域环境因子相关显著性检验
注:**表示相关性具高度统计意义(< 0.01);*表示相关性具统计意义(< 0.05)。
Note: **denotes that the correlation is highly significant with an alpha level of 0.01(2-tailed), and*denotes that the correlation is significant with an alpha level of 0.05(2-tailed).
3 讨论
3.1 种类组成及优势种分析
海水温度、盐度等环境因子是影响鱼类种类组成及分布的重要因素[15-17]。两个航次均采用底拖网进行,共捕获鱼类72种,以暖水性和暖温性种类为主,具明显的亚热带区系特征,但两季的种类组成存在明显差异。秋季渔获种类数比春季的渔获种类数多15种,原因可能是鱼类对环境变化做出的响应,特别是受海水温度影响[18]。春季的调查月份为3月,正值冬末春初,海水平均温度仅为20.0 ℃,还处于较低的水平;而秋季的调查月份为10月,海水平均温度较高,为26.6 ℃,适宜鱼类的产卵、繁殖及洄游。一些洄游性鱼类如斑鰶()、黑尾小沙丁鱼()、金色小沙丁鱼()、裘氏小沙丁鱼()和海鳗()等暖水性鱼类在越冬后回到近岸海域生活。
种类组成及资源密度上,大鹏湾北部海域秋季调查鱼类为54种,优势种生物量为1 731 kg/km2;2015年大鹏半岛海域秋季渔业资源调查中鱼类为55种,平均生物量约为1 563 kg/km2[2];2015年秋季珠江口中华白海豚自然保护区渔业资源调查中鱼类为50种,平均生物量为1 630 kg/km2[19]。大鹏湾北部海域位于大鹏半岛的西部海域,而中华白海豚保护区处于大鹏半岛西南海域且相隔香港,三者的鱼类种类及生物量调查结果基本一致,表明近5年来整个珠江口近岸海域的鱼类资源基本处于稳定的状态,渔业资源的下降趋势得到了有效的遏制。大鹏湾北部海域春季调查鱼类为39种,优势种生物量为4 459 kg/km2;2015年春季珠江口中华白海豚自然保护区渔业资源调查中鱼类为35种,平均生物量为430 kg/km2[19],二者的鱼类种类数相差不大,但资源生物量相差很大,主要原因为大鹏湾北部海域的3个优势种日本海鰶()、褐蓝子鱼()和颈斑鰏()的生物量之和高达3 971 kg/km2,其中日本海鰶为表层洄游性鱼类,而褐蓝子鱼为低值鱼类,颈斑鰏常作为杂鱼喂食高经济价值之养殖鱼类。由于地理位置的差异,各海域在鱼类种类数上相差不大,但鱼类群落组成上有较大的差异,优势种也基本不同。
相对重要性指数可用来指示物种在群落中的数量比重和对栖息地的占有范围,是用于衡量群落中不同物种优势度最简单有效的方法[20]。两个航次出现的优势种中,颈斑鲾、日本海鰶、褐蓝子鱼和二长棘鲷均能保持相对较高的生物量和分布范围,是大鹏湾北部海域较为稳定的资源种类,虽然这些种类在不同季节的渔获量存在差异,但均为两个调查航次的优势种类,应归属于正常的季节波动;此外,此种差异也与调查的时间、天气以及环境因子的变化有关。根据春季的调查数据,发现部分成鱼如日本海鰶、褐蓝子鱼和黑斑多指马鲅已有成熟卵粒出现(实验室解剖观察),表明大鹏湾北部海域可能是这些鱼类的产卵场。鉴于此,在广东大鹏湾北部海域开展鱼类生物群落生物多样性保护工作有一定的生态意义,有利于维持该海域生态系统的平衡。
3.2 物种多样性
是反映群落结构稳定性的重要指标。群落物种越丰富,各种类个体数分布越均匀,和越高。当环境胁迫群落生态系统时,种类数和个体数均会减少[21-22]。调查结果显示,广东大鹏湾北部海域(1.77)与大鹏半岛最接近大鹏湾北部海域的Group C[2](约2.0)所处海域的很接近,鱼类资源多样性水平基本一致;而与珠江口中华白海豚自然保护区的平均水平(2.62)对比,其鱼类资源多样性水平明显较低。根据蔡立哲等[23]对的评价范围(1.00 ~ 2.00),广东大鹏湾北部海域已受到一定程度的干扰,其地理位置更靠近海岸,存在因港区建设和港口运行等工业活动造成海域水质污染干扰生态系统平衡的可能,需引起注意并加强环境保护力度。
根据平均分类差异指数(Δ+)的95%置信区间漏斗曲线,秋季大鹏湾北部海域鱼类群落Δ+理论平均值为35.97,除S2、S4、S10、S11站位外,大部分站位Δ+均分布在95%置信区间内,表明大鹏湾北部海域局部区域环境受到一定程度的干扰[24],与结果一致;秋季分类差异变异指数(∧+)的95%置信区间漏斗曲线中,大鹏湾北部海域鱼类群落∧+理论平均值为109.85,各站位均落于95%置信区间内,表明这些站位物种间的亲缘关系较均匀。春季大鹏湾北部海域鱼类群落Δ+理论平均值为37.69,∧+理论平均值为100.19,各站位均落在对应的95%置信区间漏斗曲线中,表明春季鱼类群落结构基本稳定。整体来看,整个大鹏湾北部海域鱼类群落结构基本稳定,受到轻微的人类活动及环境改变的影响。
3.3 鱼类群落与环境因子的关系
鱼类群落结构实质上是环境因子(水温、水深、盐度、溶解氧和营养盐等)通过影响鱼类对栖息地的选择及其对环境的耐受性,塑造出不同的地理分布格局[25]。研究表明,盐度、水深和水温对广东大鹏湾北部海域鱼类群落结构有着重要影响。5个环境因子共解释了45.93% ~ 51.18%的鱼类群落结构空间格局。秋、春季是一年中雨量相对丰沛的季节,调查的大鹏湾北部海域西侧为香港及诸岛,北侧为深圳市,均邻接陆地,水深相对较浅;入海径流、气温对其近岸海水盐度、温度的变化均会产生一定影响,又由于有盐田港的港口航道,水深变化也比较明显。因此,在秋、春两季,盐度、水深和水温对其鱼类群落结构有着显著的影响;在这5个环境因子中,水深和溶解氧、盐度和溶解氧的相关关系都达到了显著(< 0.05),这都表明了这些环境因子直接或间接的影响着鱼类群落结构。本研究发现,大鹏湾北部海域秋季鱼类资源密度及丰度均高于春季,秋季调查的鱼类资源密度和丰度分别是春季调查的1.49和1.45倍,与珠江口[19]、中街山列岛[21]及湄洲湾[26]海域的调查结果相似,表明春季水温相对较低,除部分地方性栖息种类之外,越冬洄游的种类未大规模洄游于该地,在春季繁殖期之后,由于鱼类个体数量有效补充及洄游鱼类的加入,导致秋季鱼类资源密度和丰度高于春季。
4 结论
(1)广东大鹏湾北部海域秋、春两季共捕获鱼类72种,隶属于10目35科59属。其中日本海鰶、二长棘鲷、褐蓝子鱼和颈斑鰏为两个季度共同优势种。
(2)盐度、水深是秋季显著影响广东大鹏湾北部鱼类分布的重要环境因子,温度为春季显著影响鱼类分布的重要环境因子。
(3)广东大鹏湾北部海域鱼类现存物种资源量秋高春低,其群落结构在秋季受一定的外界干扰,春季基本稳定。
[1] 陈丕茂, 秦传新, 袁华荣, 等. 南海柘林湾鱼类群落结构季节变动的研究[J]. 南方水产科学, 2017, 13(2): 26-35.
[2] 张露, 袁华荣, 佟飞, 等. 广东深圳大鹏半岛海域秋季渔业资源群落结构分析[J]. 安徽农业科学, 2018, 46(13): 83-87.
[3] 余景, 毛江美, 袁华荣, 等. 深圳鹅公湾水域渔业资源季节变动分析[J]. 南方农业学报, 2017, 48(6): 1106-1112.
[4] 国家海洋局. GB/T12763.6 海洋调查规范[S]. 北京: 中国标准出版社, 2007.
[5] 孙典荣, 陈铮. 南海鱼类检索. 上册[M]. 连云港: 海洋出版社, 2013.
[6] MARGALEF R. Information theory in ecology[J]. General Systems, 1958, 3(1): 36-71.
[7] SHANNON C E. A mathematical theory of communication[J]. Bell System Technical Journal, 1948, 27(4): 623-656.
[8] PIELOU E C. Ecological Diversity[M]. New York: John Wiley and Sons, 1975.
[9] CLARKE K R. Non-parametric multivariate analyses of changes in community structure[J]. Austral Ecology, 1993, 18(1): 117-143.
[10] WARWICK R M, TURK S M. Predicting climate change effects on marine biodiversity: comparison of recent and fossil molluscan death assemblages[J]. Journal of the Marine Biological Association of the United Kingdom, 2002, 82(5): 847-850.
[11] PINKAS L, OLIPHANT, IVERSON I L K. Food habits of albacore, bluefin tuna, and bonito in California waters[J]. Fishery Bulletin, 1971, 152: 5-10.
[12] 程济生, 俞连福. 黄、东海冬季底层鱼类群落结构及多样性变化[J]. 水产学报, 2004, 28(1): 29-34.
[13] 刘鸿, 牛建功, 刘春池, 等. 新疆额敏河干流鱼类群落结构及其与环境因子的关系[J]. 生态学杂志, 2017, 36(12): 3558-3563.
[14] 曾嘉维, 林坤, 王学锋, 等. 雷州湾附近海域鱼类群落结构及其与环境因子的关系[J]. 中国水产科学, 2019, 26(1): 108-117.
[15] 郑亮. 黄河口海域鱼类群落结构初步研究[D]. 上海: 上海海洋大学, 2014.
[16] ZHAO S J, LÜ B, LI R W, et al. A preliminary analysis of fishery resource exhaustion in the context of biodiversity decline[J]. Science China Earth Sciences, 2016, 59(2): 223-235.
[17] 邱永松. 南海北部大陆架鱼类群落的区域性变化[J]. 水产学报, 1988, 12(4): 303-313.
[18] 张洪亮, 潘国良, 王伟定, 等. 岱衢洋拖网甲壳动物多样性的季节变化[J]. 海洋与湖沼, 2012, 43(1): 95-99.
[19] 黄吉万, 孙典荣, 刘岩, 等. 珠江口中华白海豚自然保护区鱼类群落多样性分析[J]. 南方农业学报, 2018, 49(5): 1000-1007.
[20] 徐开达, 卢衎尔, 卢占晖, 等. 韭山列岛自然保护区虾类优势种生态位[J]. 生物多样性, 2018, 26(6): 601-610.
[21] 徐开达, 张洪亮, 谢汉阳, 等. 中街山列岛水域甲壳类资源及其群落多样性[J]. 海洋渔业, 2012, 34(3): 308-315.
[22] 张静, 陈永俊, 张然, 等. 2008年夏季东山湾游泳动物种类的组成和多样性[J]. 应用海洋学学报, 2013, 32(2): 222-230.
[23] 蔡立哲, 马丽, 高阳, 等. 海洋底栖动物多样性指数污染程度评价标准的分析[J]. 厦门大学学报(自然科学版), 2002, 41(5): 641-646.
[24] 朱晓芬, 陈彬, 俞炜炜, 等. 厦门湾大型底栖动物分类学多样性指数及分类充分性[J]. 生态学报, 2018, 38(15): 5554-5565.
[25] 王雪辉, 邱永松, 杜飞雁, 等. 北部湾鱼类群落格局及其与环境因子的关系[J]. 水产学报, 2010, 34(10): 1579-1586.
[26] 杨杰青, 史赟荣, 沈新强, 等. 湄洲湾海域甲壳动物群落多样性分析[J]. 南方水产科学, 2017, 13(5): 16-24.
Analysis of the Fish Community Structure and Its Relationship with Environmental Factors in Autumn and Spring in Northern Dapeng Bay, Guangdong
ZHANG Jing1,2, QIU Jun-wen1, CHEN Chun-liang3, ZENG Jia-wei1,2, WANG Xue-feng1,2
(1.,524088,; 2.,524025,; 3.,,424088,)
To provide basic data for the current status of resources, the changes of dominant populations and the stock enhancement, the fish community structure in Northern Dapeng Bay and its relationship with different environmental factors were studied.Fish community indexes were calculated and canonical correspondence analysis (CCA) was used to study the relationship between fish community structure and environmental factors according to the survey data carried out from October, 2018(autumn) to March, 2019(spring).The results showed that there were 72 fish species that belonged to 10 orders, 35 families and 59 genera.,,andwere the common dominant species in the two seasons. The Margalaf index(), Pielous evenness index() and Shannons diversity index() in autumn were higher than those in spring. The Average TD(Δ+) in autumn was 35.97, where the survey stations S2, S4, S10 and S11 were lower than the lower boundary of the 95% probability funnel, and the Variation in TD(∧+) was 109.85 which indicated a certain external disturbance on the community. TheΔ+in spring was 37.69 and the ∧+was 100.19 which indicated a certain stability on the community. CCA showed that salinity and depth were important environmental factors that significantly affected the fish distribution in autumn,which of2were 0.75 and 0.70, and the temperature was an important environmental factor that significantly affected the fish distribution in spring(2= 0.72). In autumn, the fish community structure was greatly affected by the changes in environmental factors, and the first and second axes explained to 13.32% and 12.44% of the species distribution respectively. In spring, the first and second axes explained 16.91% and 10.93% respectively.The current fish biomass in autumn is higher than that in spring in Northern Dapeng Bay, and the community structure is affected by an external disturbance in autumn and stable in spring.
Northern Dapeng Bay; fish; diversity; community structure; canonical correspondence analysis(CCA)
S931
A
1673-9159(2020)06-0043-10
10.3969/j.issn.1673-9159.2020.06.009
张静,邱俊文,陈春亮,等. 广东大鹏湾北部海域春秋季鱼类群落结构及其与环境因子关系[J]. 广东海洋大学学报,2020,40(6):43-52.
2020-07-07
广东省大学生创新创业训练计划项目(230419179002);南方海洋科学与工程广东省实验室(湛江)资助项目(ZJW-2019-06)
张静(1977-),女,博士,副教授,从事海洋环境生态学的科研与教学工作。E-mail:zjouzj@126.com
王学锋,教授,研究方向为渔业生态与环境。E-mail:xuefeng1999@126.com
(责任编辑:刘朏)
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
儿童时代·幸福宝宝(2020年9期)2020-09-08
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
摄影之友(影像视觉)(2017年1期)2017-07-18
小学阅读指南·低年级版(2016年6期)2016-05-14
