
鲍鱼肠多肽对LPS诱导HUVEC炎症的抑制作用
2020-10-27 08:59:34萧振邦黄晓洋陈佳丽杨胜涛千忠吉
广东海洋大学学报 2020年6期
萧振邦,黄晓洋,陈佳丽,杨胜涛,刘 怡,千忠吉,3
鲍鱼肠多肽对LPS诱导HUVEC炎症的抑制作用
萧振邦1,黄晓洋1,陈佳丽2,3,杨胜涛2,刘 怡1,千忠吉1,3
(1. 广东海洋大学化学与环境学院,广东 湛江 524088;2. 广东海洋大学食品科技学院,广东 湛江 524088;3. 广东海洋大学深圳研究院,广东 深圳 518108)
【】探究序列为KVEPQDPSEW的鲍鱼肠多肽AATP对LPS诱导HUVEC人脐静脉内皮细胞的抗炎作用。用“MTT”法检测细胞活力,“DCFH-DA”法测定活性氧(ROS)的含量,用蛋白免疫印迹法检测细胞间黏附分子-1(ICAM-1)、血管细胞黏附分子-1(VCAM-1)和内皮素-1(ET-1)以及NF-κB信号通路相关蛋白的表达情况,酶联免疫实验检测白细胞介素-6(IL-6)的释放量,分子对接分析AATP与Toll样受体(TLR4)蛋白的相互作用。“MMT”法证明AATP对HUVEC细胞无明显毒性作用(> 0.05),且能提高脂多糖(LPS)诱导组的HUVEC细胞活力(< 0.05);与对照组相比,随着实验组AATP浓度增加,ROS含量明显逐渐减少(< 0.05),内皮炎症因子ICAM-1、VCAM-1、ET-1和IL-6的表达也明显逐渐减少(< 0.05),NF-κB信号通路中p65、I-κBα的磷酸化表达显著下降(< 0.05);对接结果表明,AATP与TLR4能形成稳定的氢键,有相互作用的可能性。AATP表现出很好的清除ROS作用,可明显抑制HUVEC的炎症因子的表达和NF-κB信号通路的激活。
鲍鱼;多肽;HUVEC;抗炎;脂多糖
当前全世界心血管疾病的死亡率居高不下[1],其中动脉粥样硬化是触发心血管疾病的关键因素[2]。动脉粥样硬化的发病机制复杂,通常表现为血管炎症、内皮功能障碍、胆固醇积聚等病理特征[3]。开发出针对炎症反应和内皮功能障碍的药物降低死亡率,已成为当下研究的热点。研究发现,细菌感染引起的炎症与动脉粥样硬化间有着紧密联系[4]。由于传统抗生素直接治疗动脉粥样硬化的效果并不理想,寻找非抗生素药物来代替传统抗生素药物,成为动脉粥样硬化治疗的新研究方向。肽和肽类药物调节先天免疫系统是一个自然调节的过程,用其来控制人体对细菌感染的基本先天反应,可以最大限度地降低发生耐药性的风险。这种使用肽和肽类药物来增强抗感染反应的免疫调节同时减少有害的促炎反应的治疗方式已成为研究学者关注的方向[5]。大量研究表明,许多肽和肽类药物显示出良好的活性,如在高血压等心血管疾病方面[6],尤其是在LPS引起的炎症反应方面[7]。
鲍鱼作为一种含有优良蛋白质、脂肪酸和多糖的海生腹足动物,有海洋“软黄金”的美称[8]。研究发现,从鲍鱼中提取的活性物质具有抗癌、抗菌和抗炎的能力[9-11]。多肽Lys-Val-Glu-Pro-Glu-Asp- Pro-Ser-Glu-Trp (AATP)是近期从皱纹盘鲍(Hannai)肠道中提取纯化出的一种新型多肽,已发现AATP具有抑制癌细胞迁移和抑制血管生成的抗肿瘤作用[12],表明AATP有较好的抗癌能力。而AATP作为从鲍鱼肠道中提取的活性物质,其抑制炎症反应的作用尚未见报道。因此,本研究以活性氧(ROS)的清除效果和相关炎症因子的表达以及NF-κB信号通路活化情况来研究AATP的抗炎作用,并通过分子对接的方法预测AATP与TLR4的作用机制,从而揭示AATP在脂多糖(LPS)促炎过程中的影响,以期为鲍鱼肠多肽应用在海洋生物活性成分研究上提供科学依据。
1 材料与方法
1.1 材料
1.1.1 试剂与细胞株 人脐静脉内皮细胞HUVEC购于苏州北纳创联生物技术有限公司;鲍鱼肠多肽AATP(KVEPQDPSEW,1 214.30 u)在本课题组之前的研究中获得[12],保存温度≤-20 ℃;DMEM高糖培养基、胰蛋白酶、青霉素/链霉素(双抗)、胎牛血清FBS购于美国Gibco公司;四甲基偶氮唑盐(MTT)购于上海西格玛奥德里奇贸易有限公司;BCA蛋白试剂盒、预染蛋白marker购于上海赛默飞世尔科技有限公司;脂多糖LPS、2,7-二氯荧光黄双乙酸盐(DCFH-DA)荧光探针、细胞裂解液、PMSF、5X蛋白上样缓冲液等购于上海碧云天生物技术有限公司;一抗鼠源ICAM-1、VCAM-1、ET-1、p-p65、p65、p-I-κBα、I-κBα、二抗羊抗鼠IgG购于Santa Cruz生物技术公司;人IL-6 ELISA试剂盒购于深圳欣博盛生物科技有限公司;ECL化学发光液和0.22 μm硝酸纤维素膜NC膜购于GE Amersham公司。
1.1.2 仪器和设备 二氧化碳培养箱,松下健康医疗器械株式会社;酶标仪,Bioteck仪器公司;倒置荧光显微镜,日本东京奥林巴斯有限公司;小型垂直电泳槽、电转槽,Bio-Rad公司;全自动化学发光成像分析系统,上海天能科技有限公司。
1.2 方法
1.2.1 细胞培养 HUVEC细胞放在体积分数为1%双抗和体积分数10% FBS的DMEM中于37 ℃、体积分数5% CO2气体的培养箱中培养。对细胞设计实验分组:空白组(细胞+培养基)、对照组(细胞+培养基+100 ng/mL LPS)、实验组(细胞+培养基+100 ng/mL LPS+不同浓度AATP)。
1.2.2 样品毒性检测 HUVEC细胞在96孔板(4×105mL-1,100 µL)中培养24 h。随后加入新鲜的无血清培养基和AATP(10、30、50 μmol/L)孵育24 h。移去培养基,并向每个孔中加入100 µL MTT溶液(1 mg/mL),然后放入37 ℃培养箱中避光孵育4 h。移去MTT,并加入100 µL DMSO充分振荡溶解甲瓒晶体。使用酶标仪测定540 nm处的光密度。
1.2.3 AATP对LPS诱导HUVEC的保护作用 根据陈佳丽等的[13]方法,通过“MTT”法评估测定细胞毒性。HUVEC细胞在96孔板(4×104mL-1,100 µL)中培养24 h。随后加入新鲜的无血清培养基和AATP(10、50 μmol/L)预处理1 h后,加入LPS(100 ng/mL)孵育24 h,LPS诱导浓度参考了Mancilla-Herrera等[14]方法。移去培养基,并向每个孔中加入100 µL MTT溶液(1 mg/mL),然后放入37 ℃培养箱中避光孵育4 h。移去MTT,并加入100 µL DMSO充分振荡溶解甲瓒晶体。使用酶标仪测定540 nm处的光密度。
1.2.4 ROS的测定 通过测定DCFH-DA的荧光强度来测定HUVEC中ROS的产生。将HUVEC接种于24孔板(3×105mL-1)中,用不同浓度ATTP(10、50 μmol/L)预处理1 h,然后用LPS(100 ng/mL)孵育24 h。随后用磷酸盐缓冲液(PBS)清洗细胞3次,加荧光探针(每孔500 µL 10 μmol/L DCFH-DA)后,在培养箱中避光孵育30 min。用荧光倒置显微镜拍照,并用Image J软件分析。
1.2.5 Western blot法检测相关蛋白表达 HUVEC细胞在6孔板(5×106mL-1)中培养。用AATP(10、50 μmol/L)预处理细胞1 h,然后用LPS(100 ng/mL)处理细胞24 h。用4 ℃预冷PBS洗涤HUVEC细胞,然后在冰上用溶解缓冲液溶解30 min,用Pierce-BCA蛋白定量试剂盒测定蛋白浓度。等量蛋白质(20 ~ 40 μg)用体积分数10% SDS-PAGE进行分离,然后转移到硝化纤维NC膜上。在室温下,加入质量分数5%脱脂牛奶在TBST中封闭膜。4 h后,用一抗(稀释比例为1∶500)在4 ℃下孵育过夜。在室温下用二抗(稀释比例为1∶4 000)孵育2 h,然后用TBST洗涤4次。最后,使用增强化学发光检测系统对膜进行显色观察并拍照记录。
1.2.6 酶联免疫分析 HUVEC细胞在6孔板(5×106mL-1)中培养。用AATP(10、50 μmol/L)预处理细胞1 h,然后用LPS(100 ng/mL)处理细胞24 h。24 h后12 000 r/min离心10 min后提取上清液。根据酶联免疫试剂盒说明书进行IL-6含量测定。
分别设空白孔、标准孔和待测样品孔,取出实验所需孔板,每孔加入100 μL的不同浓度上清液和不同浓度标准品,在37 ℃培养箱中孵育90 min;洗板5次,甩尽孔中液体后,加入100 μL的生物素抗体工作液,在37 ℃孵育60 min;洗板5次,甩尽孔中液体后,加入100 μL的酶结合物工作液,在37 ℃避光孵育30 min;洗板5次,甩尽孔中液体后,每孔加100 μL显色液,37 ℃避光孵育15 min;最后每孔加入100 μL终止液终止反应,蓝色立转黄色,混匀后立刻利用酶标仪检测在波长为450 nm处的光密度。根据测量数据制作标准曲线,并根据标准曲线计算IL-6的分泌量。
1.2.7 分子对接模拟 从蛋白质数据库(http://www.rcsb.org/PDB/)下载TLR4(PDB代码:2Z64)的三维晶体结构。为了模拟受体活性位点与配体分子的相互作用,使用了Discovery Studio 3.5中的CDOCKER算法。采用高分子动力学方法随机搜索小分子(AATP)的构象,并采用模拟退火方法优化受体活性位点(TLR4)的构象,进行CDOCKER对接研究。
1.3 统计学分析
所有的实验结果都是进行3次平行实验分析得出的,并用平均值±SD表示。采用SPSS statistics 25进行统计学处理,Image J、Graphpad Grism 5.0软件进行分析制图。所有数据均采用单因素方差分析(ANOVA)和Dunnett多重比较检验进行组间比较,显著性水平为为0.05。
2 结果与分析
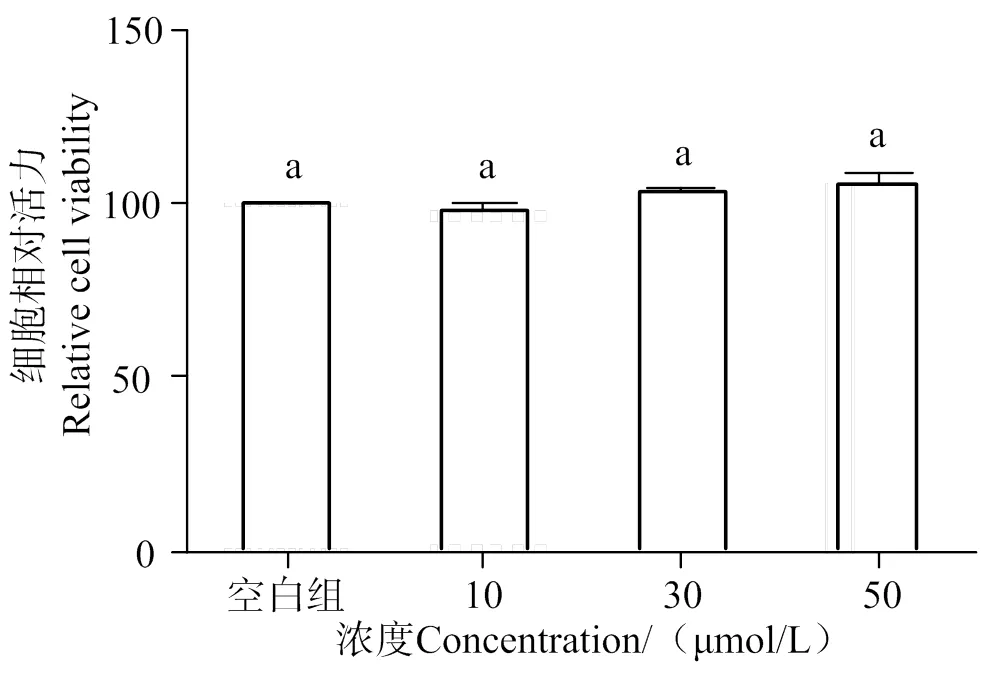
2.1 细胞毒性实验
如图1所示,不同浓度处理组的AATP对HUVEC细胞活力无明显影响(> 0.05),这说明AATP在10 ~ 50 μmol/L范围内对HUVEC细胞无毒性作用,可选用该范围的浓度进行后续实验。
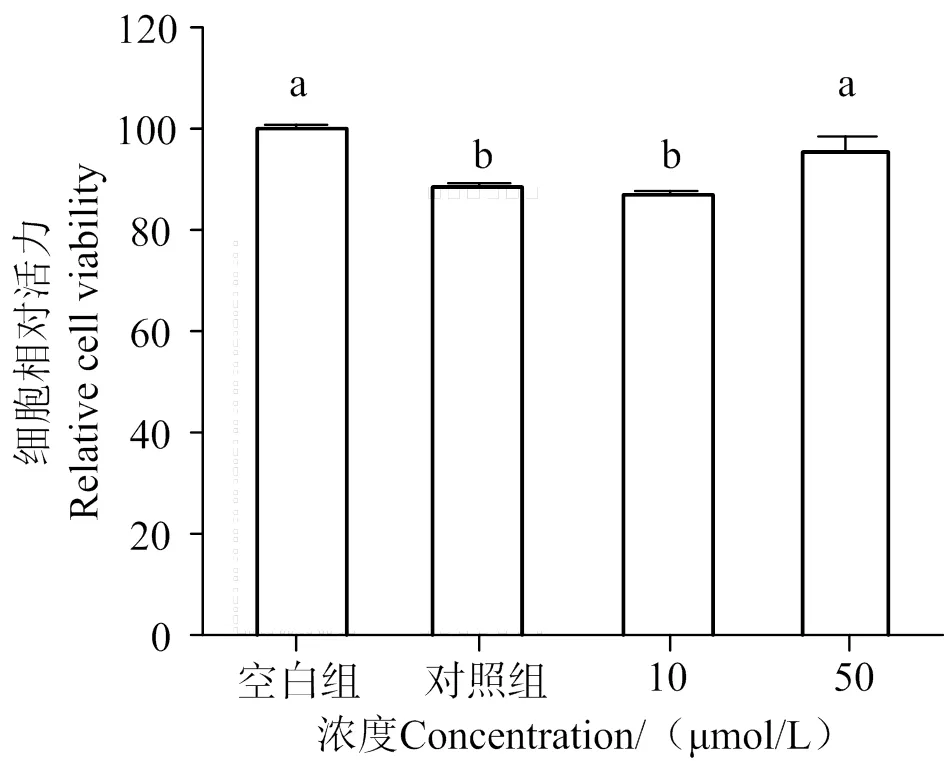
2.2 AATP对LPS诱导的HUVEC细胞的保护作用
如图2所示,与空白组相比,LPS的刺激明显降低细胞活力(< 0.05)。与对照组相比,50 μmol/L以上的AATP可使细胞相对活力提高(< 0.05)。实验结果表明,在50 μmol/L浓度范围内,AATP对LPS诱导的HUVEC细胞表现了保护作用。
凡含一个相同字母表示差异不显著(P > 0.05)
凡含一个相同字母表示差异不显著(P > 0.05)
2.3 AATP对LPS诱导的HUVEC细胞的ROS清除作用
如图3、4所示,与空白组相比,对照组LPS诱导后的ROS水平有明显升高(< 0.05)。AATP处理保护的样品组ROS水平降低。显然,AATP以剂量依赖的方式显著抑制ROS的产生。结果发现,10 μmol/L AATP可有效降低LPS诱导的ROS生成(< 0.05)。
2.4 AATP对LPS诱导后HUVEC黏附因子和血管收缩因子的表达
如图5~8所示,通过分析各组条带灰度可知,与空白组相比,LPS的刺激明显增加了黏附因子ICAM-1、VCAM-1和血管收缩因子ET-1的表达(< 0.05),随着10、50 μmol/L AATP浓度处理,其表达有显著性下调(< 0.05)。

图3 不同浓度AATP对HUVEC细胞的清除ROS作用

凡含一个相同字母表示差异不显著(P > 0.05)

图5 LPS诱导后HUVEC细胞ICAM-1、VCAM-1、ET-1的表达

凡含一个相同字母表示差异不显著(P > 0.05)

凡含一个相同字母表示差异不显著(P > 0.05)

凡含一个相同字母表示差异不显著(P > 0.05)
2.5 AATP对LPS诱导后NF-κB信号通路的阻断作用
如图9~11所示,通过分析各组条带灰度可知,与空白组相比,LPS的刺激明显提高了p-p65、p-I-κBα的水平(< 0.05),随着10、50 μmol/L AATP浓度处理,其表达有显著性下调(< 0.05)。这表明AATP可以通过抑制p65、I-κBα的磷酸化从而达到阻断NF-κB信号通路的作用。

图9 LPS诱导后HUVEC细胞p-p65、p65、p-I-κBα、I-κBα的表达情况

凡含一个相同字母表示差异不显著(P > 0.05)

凡含一个相同字母表示差异不显著(P > 0.05)
2.6 AATP对IL-6的抑制作用
通过酶联免疫实验测定AATP对IL-6分泌量的影响。如图12所示,与空白组相比,加入LPS的对照组的IL-6分泌量有显著增加(< 0.05)。与对照组相比,加入50 μmol/L AATP HUVEC细胞的IL-6分泌量显著下降(< 0.05)。说明在LPS的刺激下,AATP具有下调HUVEC细胞IL-6分泌量的作用。
2.7 AATP与TLR4的相互作用
利用分子对接检测AATP与TLR4配体之间的相互作用。通过模拟分子对接Discovery Studio软件得到AATP与TLR4配体的可能结合位点,取结合能最小值的构象作为最佳构象结果。图13a所示关于AATP-TLR-4对接的CDOCKER结果结合能为-201.64 kJ/mol。如图13b,AATP与TLR4的Asp403和Arg380产生2个氢键,键长为1.86×10-10m和2.75×10-10m。

凡含一个相同字母表示差异不显著(P > 0.05)
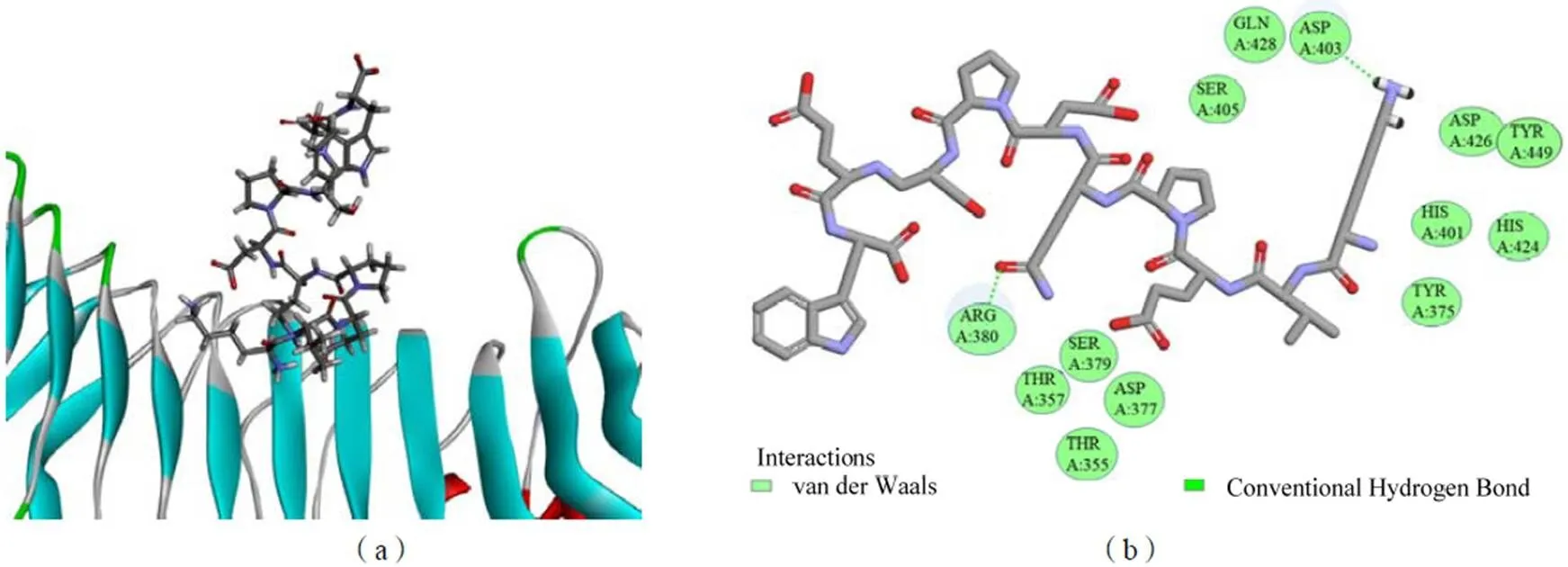
图13 AATP与TLR4的分子对接三维图(a)和二维图(b)
3 讨论
动脉粥样硬化是一种多因素的炎症性疾病,其发生与内皮功能障碍有关,而TLR-4牵涉其中这些炎症过程,包括损伤性自由基的形成、内皮功能障碍和促炎因子过度表达[15]。大量的研究表明,LPS可以直接作用于HUVEC,使其发生长梭状变形,并引起细胞黏附因子的分泌和细胞凋亡,这个过程与TLR-4识别LPS后高度表达有密切关系[16-18]。近年来肽和肽的化合物的抗炎活性越来越受关注,例如从马蹄蟹()血细胞中提取的TP-1(KWCFRVCYRGICYRRCR-NH2)[19]。由于多肽具有较高的生物活性和易于合成等特点,在临床试验中得到了广泛的应用。本研究发现鲍鱼肠多肽AATP(KVEPQDPSEW)对LPS诱导的HUVEC具有良好的抗炎作用,这说明AATP有成为抗炎药物的潜力。
之前内皮功能障碍的研究中表明,LPS诱导的炎症与氧化应激有关[20-21]。细胞内过量ROS可被视为氧化应激的原因之一,ROS可促进动脉粥样硬化发展过程中内皮细胞凋亡[7]。此外,ROS还可以促进促炎因子的表达,进而导致炎症反应和内皮功能障碍[22]。AATP对LPS诱导HUVEC细胞的ROS清除作用表明AATP具有降低ROS含量的作用,减少内皮功能障碍发生的可能[23]。
在炎症起始阶段,ICAM-1和VCAM-1黏附分子的大量产生是炎症发生的重要标志。黏附分子与白细胞结合后,刺激内皮细胞改变形态,细胞间隙可作为白细胞的迁移通道。降低黏附分子的表达可有效降低血管内皮细胞的通透性[24]。本研究发现,AATP可以明显下调LPS诱导后HUVEC中ICAM-1、VCAM-1、ET-1和IL-6的蛋白含量,这些都是参与白细胞黏附和调节血管收缩的炎症因子。这预示着具有阻断作用的AATP不仅具有抗动脉粥样硬化作用,而且具有抗炎作用。HUVEC是ET-1的主要生物来源[25],ET-1的大量增加可能预示着血管收缩功能的失衡并导致血管损伤,引起高血压从而让动脉粥样硬化发生和发展[26],此外LPS预处理的HUVEC中ET-1的增加也可能进一步促进ROS的生成[23]。
TLR4作为主要识别LPS的受体,在炎症反应中起着重要作用,与动脉粥样硬化和高血压等心血管疾病有着密切关系[27]。TLR4识别LPS后,可以激活细胞内的下游信号通路NF-κB并且诱导内皮细胞相关的ICAM-1、VCAM-1、IL-6、IL-8等炎症因子的生成[28],AATP 通过抑制p65、I-κBα的磷酸化对NF-κB信号通路的阻断说明AATP在HUVEC细胞中可以发挥作用。分子对接的结果表明,AATP与TLR4产生两个稳定的氢键,可能存在对接位点,具有相互作用的可能性。这说明AATP可能通过与TLR4相互作用的方式,阻断TLR4-NF-κB通路,有效减少黏附分子、炎症因子的表达,
本研究表明,AATP是具有一定的抗炎作用,通过抑制促炎因子ROS的过表达,炎症相关的NF-κB信号通路,发挥抗炎作用。同时可以使内皮炎症因子ICAM-1、VCAM-1、ET-1和IL-6含量下调,是一种潜在的抑制炎症反应的活性物质。但本研究仅从蛋白水平对相关炎症因子进行探讨,对基因水平的研究尚不完善,今后的研究可以通过检验相关炎症因子的mRNA的变化并结合相关蛋白水平的研究,去进一步验证AATP的抗炎活性。
4 结论
本研究证明体外实验中AATP能够明显减少HUVEC的ROS生成,表现出很好的清除ROS能力以减缓细胞损伤,AATP可降低细胞相关炎症因子的表达,抑制NF-κB信号通路相关蛋白的表达。分子对接结果表明AATP和TLR4之间能形成稳定氢键,存在相互作用的可能性。这些结果表明AATP具有成为潜在抗炎活性物质的可能性,并有助于为海洋生物多肽的活性研究提供有效佐证。
[1] WENS I, DALGAS U, STENAGER E, et al. Risk factors related to cardiovascular diseases and the metabolic syndrome in multiple sclerosis - a systematic review[J]. Multiple Sclerosis (Houndmills, Basingstoke, England), 2013, 19(12): 1556-1564.
[2] MERKIN S S, KARLAMANGLA A, ELASHOFF D, et al. Change in cardiometabolic score and incidence of cardiovascular disease: the multi-ethnic study of atherosclerosis[J]. Annals of Epidemiology, 2015, 25(12): 912-917.
[3] LIBBY P, HANSSON G K. Inflammation and immunity in diseases of the arterial tree[J]. Circulation Research, 2015, 116(2): 307-311.
[4] 朱建健, 王宪. 慢性炎症、自身免疫和动脉粥样硬化[J]. 生理科学进展, 2002, 33(4): 327-331.
[5] SKOVBAKKE S L, FRANZYK H. Anti-inflammatory properties of antimicrobial peptides and peptidomimetics: LPS and LTA neutralization[J]. Antimicrobial Peptides, 2017, 1548: 369-386.
[6] ZHU Z S, LI J Y, ZHANG X R. Astragaloside IV protects against oxidized low-density lipoprotein (ox-LDL)-induced endothelial cell injury by reducing oxidative stress and inflammation[J]. Medical Science Monitor, 2019, 25: 2132-2140.
[7] 刘旭峰, 闵旭晨, 谷雪芳, 等. 抗感染治疗与动脉粥样硬化之间的关系[J]. 中国动脉硬化杂志, 2020, 28(5): 452-455.
[8] 李帅鹏, 晁珊珊, 高仕林. 我国鲍鱼养殖产业现状与对策[J]. 江西水产科技, 2019(6): 44-46.
[9] DE ZOYSA M, NIKAPITIYA C, WHANG I, et al. Abhisin: a potential antimicrobial peptide derived from histone H2A of disk abalone ()[J]. Fish & Shellfish Immunology, 2009, 27(5): 639-646.
[10] QIAN Z J, BOMI R, WON S P, et al. Inhibitory effects and molecular mechanism of an anti-inflammatory peptide isolated from intestine of abalone, haliotis discus hannai on LPS-induced cytokine production via the p-p38/p-JNK pathways in RAW264.7 macrophages[J]. Journal of Food and Nutrition Research, 2016, 4(10): 690-698.
[11] NGUYEN V, QIAN Z J, JUNG W. AbaloneHannai intestine digests with different molecule weights inhibit MMP-2 and MMP-9 expression in human fibrosarcoma cells[J]. Fisheries and Aquatic Sciences, 2012, 15(2): 137-143.
[12] GONG F, CHEN M F, ZHANG Y Y, et al. A novel peptide from abalone (Hannai) to suppress metastasis and vasculogenic mimicry of tumor cells and enhance anti-tumor effect in vitro[J]. Marine Drugs, 2019, 17(4): 244.
[13] 陈佳丽,杨胜涛,萧振邦,等. 罗非鱼鱼皮肽的体外ACE抑制活性、消化性及分子对接对比[J]. 广东海洋大学学报, 2019, 39(5): 107-114.
[14] MANCILLA-HERRERA I, ALVARADO-MORENO J A, CÉRBULO-VÁZQUEZ A, et al. Activated endothelial cells limit inflammatory response, but increase chemoattractant potential and bacterial clearance by human monocytes[J]. Cell Biology International, 2015, 39(6):721-732.
[15] BOTHAM K M, WHEELER-JONES C P D. Postprandial lipoproteins and the molecular regulation of vascular homeostasis[J]. Progress in Lipid Research, 2013, 52(4): 446-464.
[16] 张旭, 许冬青, 朱平, 等.LPS诱导HUVEC凋亡的分子机制[J]. 中医药学刊, 2006(12): 2211-2213.
[17] 程君涛, 袁建成, 郑江, 等. 内毒素对人脐静脉内皮细胞形态和功能的影响[J]. 中华烧伤杂志, 2001, 17(3): 155-158.
[18] 李真玉. TLR4基因沉默对兔颈动脉粥样硬化斑块血管生及细胞凋亡的影响及机制探讨[D]. 天津: 天津医科大学, 2015.
[19] SARAVANAN R, MOHANRAM H, JOSHI M, et al. Structure, activity and interactions of the cysteine deleted analog of tachyplesin-1 with lipopolysaccharide micelle: Mechanistic insights into outer-membrane permeabilization and endotoxin neutralization[J]. Biochimica et Biophysica Acta, 2012, 1818(7): 1613-1624.
[20] GUZIK T J, WEST N E, BLACK E, et al. Vascular superoxide production by NAD(P)H oxidase: association with endothelial dysfunction and clinical risk factors[J]. Circulation Research, 2000, 86(9): E85-E90.
[21] ZHANG M, PAN H C, XU Y J, et al. Allicin decreases lipopolysaccharide-induced oxidative stress and inflammation in human umbilical vein endothelial cells through suppression of mitochondrial dysfunction and activation of[J]. Cellular Physiology and Biochemistry, 2017, 41(6): 2255-2267.
[22] MURDOCH C E, CHAUBEY S, ZENG L F, et al. Endothelial NADPH oxidase-2 promotes interstitial cardiac fibrosis and diastolic dysfunction through proinflammatory effects and endothelial-mesenchymal transition[J]. Journal of the American College of Cardiology, 2014, 63(24): 2734-2741.
[23] GOPALAKRISHNA D, PENNINGTON S, KARAA A, et al. ET-1 stimulates superoxide production byfollowing exposure of vascular endothelial cells to endotoxin[J]. Shock (Augusta, Ga.), 2016, 46(1): 60-66.
[24] COOK-MILLS J M, MARCHESE M E, ABDALA-VALENCIA H. Vascular cell adhesion molecule-1 expression and signaling during disease: regulation by reactive oxygen species and antioxidants[J]. Antioxidants & Redox Signaling, 2011, 15(6): 1607-1638.
[25] YANAGISAWA M, KURIHARA H, KIMURA S, et al. A novel potent vasoconstrictor peptide produced by vascular endothelial cells[J]. Nature, 1988, 332(6163): 411-415.
[26] CHEN J L, GONG F, CHEN M F, et al. In vitro vascular-protective effects of a tilapia by-product oligopeptide on angiotensin II-induced hypertensive endothelial injury in HUVEC by Nrf2/NF-κB pathways[J]. Marine Drugs, 2019, 17(7): 431.
[27] HERNANZ R, MARTÍNEZ-REVELLES S, PALACIOS R, et al. Toll-like receptor 4 contributes to vascular remodelling and endothelial dysfunction in angiotensin II-induced hypertension[J]. British Journal of Pharmacology, 2015, 172(12):3159-3176.
[28] DAYANG E Z, PLANTINGA J, TER ELLEN B, et al. Identification of LPS-activated endothelial subpopulations with distinct inflammatory phenotypes and regulatory signaling mechanisms[J]. Frontiers in Immunology, 2019, 10: 1169.
Effect of Abalone Polypeptide on LPS Induced HUVEC Cell Inflammation
XIAO Zhen-bang1, HUANG Xiao-yang1, CHEN Jia-li2,3, YANG Sheng-tao2, LIU Yi1, QIAN Zhong-ji1,3
(1.,,524088,; 2.,,524088,; 3.,518108,)
To analyze the anti-inflammatory effect of abalone intestinal polypeptide AATP sequenced KVEPQDPSEW on human umbilical vein endothelial cells (HUVEC) induced by LPS.MTT method was used to detect cell viability; DCFH-DA was used to detect ROS content; western blotting was used to detect the expression of ICAM-1, VCAM-1, ET-1 and other proteins in the NF-κB signaling pathway, and ELISA test to detect the release of IL-6; molecular docking analysis was used to analyze the interaction between AATP and toll-like receptor (TLR4).MMT method showed that AATP had no significant toxic effect on HUVEC(> 0.05). It could improve the HUVEC viability of LPS induced group(< 0.05). Compared with the control group, AATP concentration in the experimental group increased, ROS content decreased significantly(< 0.05). The expression of ICAM-1, VCAM-1, ET-1, IL-6 and phosphorylated p65, phosphorylated I-κBα in the NF-κB signaling pathway decreased gradually(< 0.05). The docking results showed that AATP and TLR4 could form stable hydrogen bonds, which had the possibility of interaction.AATP can clear ROS and inhibit the expression of inflammatory factors and suppress the activation of NF-κB signaling pathway in HUVEC.
abalone; peptide; HUVEC; anti-inflammatory; LPS
Q71
A
1673-9159(2020)06-0108-08
10.3969/j.issn.1673-9159.2020.06.017
萧振邦,黄晓洋,陈佳丽,等. 鲍鱼肠多肽对LPS诱导HUVEC炎症的抑制作用[J]. 广东海洋大学学报,2020,40(6):108-115.
2020-06-18
广东省基础与应用基础研究基金项目(2020A1515011075)
萧振邦(1995-),男,硕士研究生,研究方向为海洋活性物质研究与开发。E-mail:btplk@sina.com。
黄晓洋(1999-),男,本科生,研究方向为海洋活性物质研究与开发。E-mail:784030908@qq.com
千忠吉(1978-),男,博士,副教授,研究方向为南海生物活性物质的研究与利用。E-mail:zjqian78@163.com
(责任编辑:刘朏)
猜你喜欢
意林彩版(2022年2期)2022-05-03 10:25:08
意林彩版(2022年4期)2022-05-03 00:07:57
中华养生保健(2020年7期)2020-11-16 01:13:32
文萃报·周二版(2019年2期)2019-09-10 07:22:44
现代园艺(2017年13期)2018-01-19 02:28:09
中成药(2017年9期)2017-12-19 13:34:20
中成药(2017年10期)2017-11-16 00:50:09
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
中药与临床(2015年5期)2015-12-17 02:39:30
药学与临床研究(2015年4期)2015-06-05 11:35:54
