
OIP5-AS1和miR-410在胶质瘤中的表达及调控机制
2020-08-26 01:01孙伟力康天孙建平王苑宇孙晓枫杨悦焦保华
河北医药 2020年15期
孙伟力 康天 孙建平 王苑宇 孙晓枫 杨悦 焦保华
脑胶质瘤是起源于神经上皮的肿瘤的统称,年发病率为(3~8)/10万。和其他肿瘤一样,胶质瘤的发病也是由于遗传易感因素和环境中的多种致癌因素相互作用的结果。越来越多的研究表明:在多种肿瘤中出现的非编码RNA的表达失调,可能参与了调控肿瘤发生和发展过程[1]。近年来,研究者对LncRNA和microRNA在胶质瘤发生和发展过程中的作用进行了大量研究。有报道证实LncRNA TUG1在胶质瘤组织中表达下调,可发挥肿瘤抑制因子的作用,TUG1在体外过表达可以促进胶质瘤细胞的凋亡,这提示TUG1可能是治疗胶质瘤的潜在治疗靶点[2]。另据报道,在胶质瘤组织中LncRNA H19的过表达与患者的预后呈负相关,沉默H19可以通过调节miR-675的表达抑制胶质瘤细胞的生长[3]。OIP5-AS1是从编码OIP5的同一基因的反义方向转录的,最初在斑马鱼中被称为cyrano,发现其在中枢神经系统的早期发育中起关键作用[4]。在胶质瘤的发生和进展过程中,OIP5-AS1是否存在调节作用,目前笔者尚未发现相关研究。已有研究报道:miR-410的表达量随胶质瘤级别的升高而降低,可以靶向作用于下游抑癌基因MET,有效调控胶质瘤细胞的增殖和侵袭[5]。在本研究中,将探讨OIP5-AS1和miR-410在脑胶质瘤组织中表达的差异性及与胶质瘤恶性程度的关系,并进一步探讨二者相互作用的机制。
1 资料与方法
1.1 一般资料 收集2016年10月至2017年10月在河北医科大学第二医院神经外科手术切除的胶质瘤患者标本65例(胶质瘤组),所有患者术前未进行其他治疗,且不合并其他系统的原发性恶性肿瘤、严重全身感染以及结缔组织疾病。收集同期10例行去骨瓣减压术的重度颅脑外伤患者术中为降低颅内压,部分切除的脑组织标本作为对照组。
1.2 仪器与试剂 Real-Time PCR System购自美国Applied Biosystem公司;Dual-Luciferase® Reporter Assay Syste购自普洛麦格生物技术有限公司;BX51荧光显微镜购自日本Olympus公司;Trizol试剂盒购自美国Invitrogen公司;SYBR Premix Ex Tap Ⅱ、逆转录试剂盒购自日本Takara公司;HEK 293T细胞系购自中国科学院;Lipofectamine 2000购自美国Invitrogen公司;质粒由上海吉凯基因科技有限公司合成。
1.3 试验方法
1.3.1 试验分组:根据标本的性质分为肿瘤组与非肿瘤对照组,根据WHO 2007年发表的中枢神经系统肿瘤病理分类将肿瘤组再分为2组:低级别胶质瘤(Ⅰ/Ⅱ级)组,高级别胶质瘤(Ⅲ/Ⅳ级)组。
1.3.2 设计及合成引物序列:通过NCBI,选择目的基因/SNP位点,查找序列,设计引物,筛选引物,优化各组分浓度,并经BLAST检查引物特异性。由生工生物工程(上海)公司合成上述引物。见表1。

表1 引物序列
1.3.3 Real-Time PCR反应:采用Trizol试剂盒及氯仿、异丙醇、乙醇、DEPC等经裂解、匀浆、离心、洗涤、溶解后从各组胶质瘤组织、正常组织的细胞中提取总RNA。经BioTek Synergy HT多功能酶标仪鉴定待测样品OD260/OD280的比值及其浓度。然后取总RNA(2 μg)在无RNA酶离心管中配置反应体系在适当条件下进行逆转录。使用SYBR Premix Ex Taq Ⅱ试剂盒、引物、cDNA等配置总体积20 μl的qRT-PCR反应体系,反应条件:起始过程95℃ 10 min;扩增过程95℃ 15 s,60℃ 1 min,总共扩增40个循环,扩增cDNA。并以U6作为miR-410的内参基因,GAPDH作为OIP5-AS1的内参基因。待测基因的相对表达量用2-ΔΔCt进行计算,其中-ΔΔCt=-(ΔCt肿瘤组-ΔCt对照组),ΔCt肿瘤组=Ct目的基因-Ct内参基因,ΔCt对照组=Ct目的基因-Ct内参基因。实验进行3次,取平均值。
1.3.4 生物信息学分析:在Starbase 2.0数据库中,根据miR-410寻找调节其表达的LncRNA,初步明确是否为OIP5-AS1。进一步使用Target Scan预测软件,确定miR-410和OIP5-AS1的结合位点。
1.3.5 HEK 293T细胞的培养及传代:①细胞复苏、培养:37℃水浴预热含10%FBS的DMEM高糖培养基,快速解冻HEK 293T细胞系、重悬细胞,然后于1 000 r/min离心5 min。弃上清,加5 ml培养基重悬细胞后,接种到25 cm2的培养瓶中,摇匀后在5% CO2孵箱中37℃恒温培养。②细胞传代:当细胞融合度长至约90%时,给细胞传代。在超净台中弃培养基,加入5 ml PBS清洗细胞后,再加入1 ml 0.25%的胰蛋白酶消化细胞。加入5 ml含10%FBS的DMEM高糖培养基终止消化。用移液器轻轻吹下瓶壁上剩余的细胞,将细胞移入离心管中,1 000 r/min离心5 min。弃上清,重悬细胞后,接种到75 cm2培养瓶中,细胞密度在1×104细胞/cm2,恒温培养。并以1∶10传代,提取处于对数生长期的HEK 293T细胞用于双荧光素酶报告基因实验。
1.3.6 双荧光素酶报告基因实验:①实验分组:将293T细胞分为NC组(转染NC序列)和miR-410 mimics组(转染miR-410模拟物)。②构建质粒:由上海吉凯基因科技有限公司合成。③转染目的细胞质粒:提取处于对数生长期的HEK 293T细胞,当细胞融合率达到80%时,按照Lipofectamine-2000试剂盒说明书分别转染NC序列和miR-410模拟物序列。④双荧光素酶检测 转染约48 h后,加入20 μl Luciferase Assay试剂,震荡混匀后立即使用酶标仪检测萤火虫荧光酶荧光值,随后加入20 μl Stop & Glo®试剂,震荡混匀后静置3 min后使用酶标仪检测海肾荧光酶荧光值,计算萤火虫荧光素酶活性与海肾荧光素酶活性的比值。实验进行3次,取平均值。
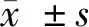
2 结果
2.1 患者临床信息及资料归纳 (1)根据标本性质分组:肿瘤组65例,其中男40例,女25例;平均年龄(46.23±12.58)岁。对照组10例,重度颅脑外伤患者。(2)根据胶质瘤病理级别分组:低级别胶质瘤组29例,其中Ⅰ级12例,Ⅱ级17例。高级别胶质瘤组36例,其中Ⅲ级19例,Ⅳ级17例。见表2。
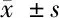
表2 5组OIP5-AS1和miR-410的表达
2.2 正常脑组织和胶质瘤组织HE染色 与正常脑组织HE染色相比,胶质母细胞瘤由分化程度低、多形性明显、胞核的非典型性突出、有丝分裂活跃的高度间变的胶质细胞组成,肿瘤的细胞密度高;肿瘤中可见明显的微血管增殖和坏死。见图1。
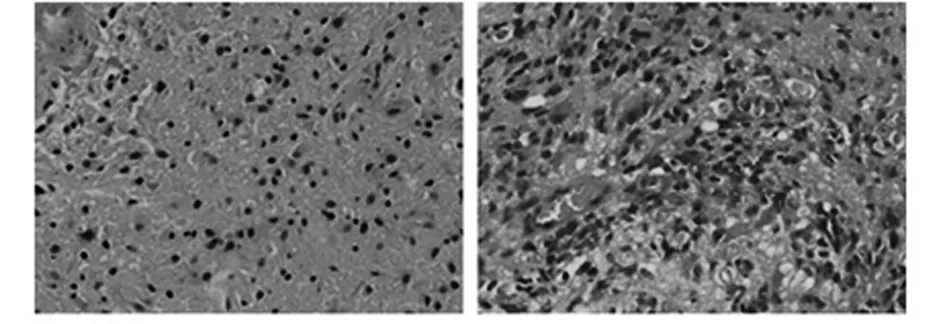
正常脑组织 胶质瘤组织(WHO Ⅳ级)
2.3 对照组与肿瘤组OIP5-AS1和miR-410的表达 肿瘤组和对照组OIP5-AS1和miR-410的表达水平。与正常脑组织比较,胶质瘤组织中OIP5-AS1的表达水平明显升高,miR-410的表达明显降低(P<0.05)。见表3,图2。
表3 对照组与肿瘤组OIP5-AS1和miR-410的表达
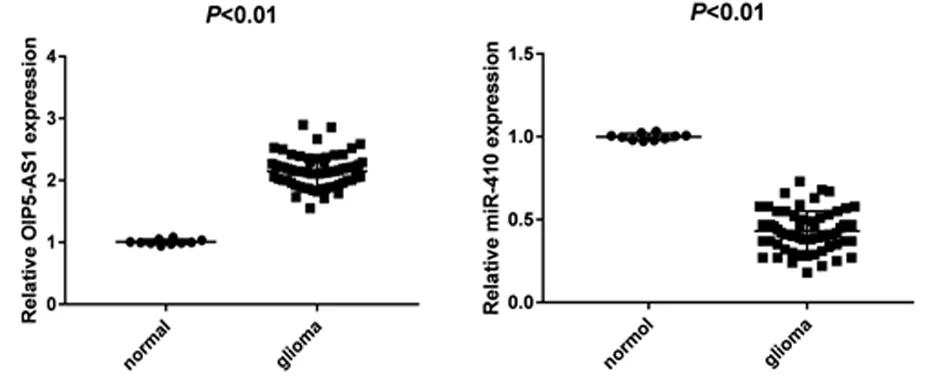
图2 OIP5-AS1和miR-410在胶质瘤和正常组织中的相对表达(散点图)
2.4 不同级别胶质瘤组OIP5-AS1和miR-410的表达水平 OIP5-AS1在高级别胶质瘤组织中的表达水平显著高于低级别胶质瘤组织(P<0.05)。miR-410在高级别胶质瘤组织中的表达水平显著低于低级别胶质瘤组织(P<0.05)。OIP5-AS1和miR-410的表达水平与胶质瘤的病理分级显著相关。见表4。
表4 不同级别胶质瘤组织OIP5-AS1和miR-410的表达
2.5 相关性分析结果 Pearson检验示:OIP5-AS1表达与miR-410表达呈负相关(r=-0.64,P<0.001)。见图3。
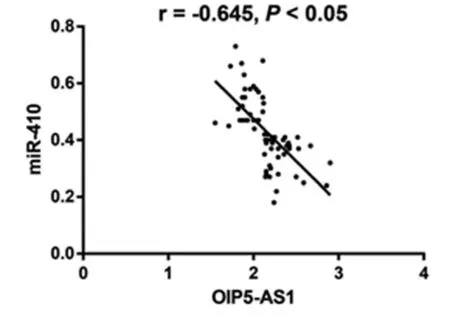
图3 OIP5-AS1表达水平与miR-410的相关性分析
2.6 OIP5-AS1与miR-410的关系 使用Starbase 2.0在线预测:OIP5-AS1是调节miR-410的分子海绵。进一步使用Target Scan预测软件确定了miR-410和OIP5-AS1可能的结合位点。见图4。

图4 在线生物信息软件预测miR-410和OIP5-AS1的结合位点
2.7 双荧光素酶活性测定结果 与NC组比较,miR-410 mimics对突变型OIP5-AS1-mt荧光素酶活性的影响无显著差异;miR-410 mimics显著下调野生型OIP5-AS1-wt的荧光素酶活性。实验结果证实:OIP5-AS1与miR-410之间存在靶向关系。见表5。
表5 双荧光素酶活性测定结果
3 讨论
胶质瘤是发病率最高的原发性颅内肿瘤,2007年世界卫生组织(WHO)中枢神经系统肿瘤分类将脑胶质瘤分为Ⅰ~Ⅳ级,其中Ⅰ、Ⅱ级为低级别脑胶质瘤,Ⅲ、Ⅳ级为高级别脑胶质瘤。在本实验中,将此作为肿瘤组的分组标准,并进一步检测了OIP5-AS1和miR-410在各组中的表达,通过观察这两种非编码RNA在不同级别胶质瘤中表达的变化趋势并经统计学证实二者与肿瘤恶性程度相关。
已有研究表明,与OIP5-AS1编码同一基因的正义方向转录的OIP5在正常组织呈限制性表达,但在肾癌及乳腺癌等许多肿瘤中过表达,因此被认为是一种致瘤基因,并且有可能成为靶向药物治疗的靶点[6]。目前笔者尚未发现相关研究表明编码OIP5的同一基因的反义方向转录的OIP5-AS1在胶质瘤发生及发展中的作用。有研究发现,OIP5-AS1在哺乳动物中普遍存在,在人类某些肿瘤中明显高表达[7]。OIP5-AS1在神经系统中表达水平较高,且在发育过程中对调控神经发育起着非常重要的作用。同时,OIP5-AS1也是近年来肿瘤相关研究的一个热点。Kim等[8]的研究表明:在人宫颈癌Hela细胞中,OIP5-AS1可能作为竞争性内源RNA与RNA结合蛋白HuR结合并使其稳定,从而抑制HuR诱导的细胞增殖,参与肿瘤细胞的调控。同样,Deng等[9]研究报道,荧光素酶检测证实miR-448是OIP5-AS1的靶基因,OIP5-AS1通过与miR-448竞争性结合,正向调控下游靶基因Bcl-2,在肺腺癌组织和细胞中高表达时,发挥致癌作用,主要是影响肺癌细胞增殖、迁移和侵袭。Yang等[10]还发现,在多发性骨髓瘤中OIP5-AS1通过调节miR-410的表达,调控其靶基因KLF10,从而介导PTEN/PI3K/AKT信号通路,促进肿瘤细胞增殖、细胞周期的进展和抑制细胞凋亡。本实验明确了OIP5-AS1和miR-410的靶向调节关系。
在胶质瘤组织中,OIP5-AS1表达显著升高,而miR-410表达降低,提示OIP5-AS1与miR-410表达水平呈负相关关系,经Pearson相关性检验也已证实二者表达量变化差异有统计学意义(P<0.05)。此外,miR-410在人胶质瘤组织中表达下调,通过下调其靶基因MET,发挥抑癌作用,对胶质瘤生长产生明显抑制作用[5]。因此,我们假设OIP5-AS1可能发挥致癌基因的作用,miR-410可能作为神经胶质瘤中的抑癌基因调节某些抑癌蛋白的表达并进一步抑制肿瘤发生和/或进展。本研究还发现OIP5-AS1和miR-410的表达与胶质瘤的病理分级密切相关。其中,胶质瘤患者病理分级越高,OIP5-AS1表达水平越高,miR-410表达水平越低,提示OIP5-AS1和miR-410与胶质瘤的发病机制和发展密切相关,可以推测OIP5-AS1对miR-410可能发挥“海绵分子”的功能。
Ganesan等[11]研究发现:OIP5-AS1过表达在人类上皮起源的肿瘤中很常见,OIP5-AS1在未分化细胞的口腔肿瘤中过表达,受其调节的6个miRNA(miR-137、miR-148a-3p、miR-30a-5p、miR-30b-5p、miR-338-3p和miR-22-3p)表达下调,OIP5-AS1对miRNA具有“海绵”的功能。OIP5-AS1表观基因组分析结果表明,通过miR-143/145的重复表达,靶向干细胞特性相关转录因子,通过共表达的NEAT1、TUG1和HOTAIR对OIP5-AS1进行正向转录调控,可能是维持肿瘤细胞干细胞特点和低分化的原因,也是预后不良的原因。在胶质瘤中,可能也存在这种调节机制,OIP5-AS1过表达,发挥其对miR-410的“海绵化”功能,并靶向抑制下游抑癌因子的表达,肿瘤细胞增殖及侵袭力增强。同时,通过OIP5-AS1正向转录调控机制,表达不断增强,进一步加速肿瘤的增殖和侵袭,导致高级别的胶质瘤中OIP5-AS1过表达。
本研究通过实验证实OIP5-AS1是胶质瘤的促癌基因,通常情况下反义lncRNA可以负向调节对应mRNA的表达,因此有必要了解OIP5在肿瘤发生中的作用,从而进一步理解OIP5-AS1做为促癌基因的机制。Freitas等[12]发现OIP5在胶质母细胞瘤中的表达率较高,可达54%。同时OIP5阳性表达组患者生存期比阴性组延长25周,统计结果提示差异有统计学意义(P<0.05)。我们可以从这一结果推断OIP5在胶质母细胞瘤起到了抑制肿瘤进展的作用。最近,Dorival等[13]在T98G GBM细胞系中对OIP5的敲除效果进行了评估。结果表明,下调OIP5可以刺激胶质瘤细胞的活力,抑制环己亚硝脲(抗癌药)诱导的细胞坏死和凋亡。OIP5在GBM细胞中的表达似乎能够增强环己亚硝脲细胞毒性作用,进一步证明该基因是GBM治疗反应的潜在治疗靶点和可能的分子生物标志物。但是国内学者通过检测92例胶质瘤组织中OIP5的表达水平,发现OIP5的阳性表达率为18.47%,阳性表达率差异无统计学意义(P>0.05)[14],能否作为胶质瘤治疗的新靶点需进一步研究。OIP5在多数肿瘤中为促癌基因,而在GBM中则相反。
随着分子生物学的发展,新一代测序技术和DNA微阵列技术极大地帮助我们在全基因组范围内识别不同癌症及其亚型的miRNA和mRNA信号。通过大量实验证实,miRNA在不同肿瘤中扮演着截然相反的角色。有研究发现:miR-410在儿童急性淋巴细胞白血病病例中的表达水平均明显增高,在疾病进展中起促进作用,在体外实验得到进一步证实[15]。miR-410在肿瘤中的作用与正、负调控相关:在某些肿瘤中负性调节BAK1、CETN3和BRD7促进肿瘤细胞增殖及侵袭,在有些肿瘤中miR-410可以有效靶向c-MET、AGTR1和SNAIL等起到抑癌作用[16]。
双荧光素酶报告基因系统将含有目的DNA的序列插入到载体工具中进行实验,在转录水平测定上游刺激对下游靶基因的影响,可以快速、灵敏、高效地检测转录因子和其靶启动子区DNA相互作用的关系,从而揭示细胞信号转导通路的分子机制。
综上所述,胶质瘤组织中OIP5-AS1的表达水平明显升高,miR-410的表达明显降低,两者可能影响胶质瘤的发生和发展。OIP5-AS1和miR-410的表达与胶质瘤的病理分级相关,而且OIP5-AS1表达与miR-410表达呈负相关,而且OIP5-AS1可以靶向调节miR-410的表达。为揭示胶质瘤发生和侵袭性生长的发病机制提供了实验证据,从而为胶质瘤的基因治疗和靶向治疗提供了理论基础。
猜你喜欢
中国现代医生(2022年19期)2022-08-25
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
昆明医科大学学报(2021年2期)2021-03-29
广东蚕业(2021年1期)2021-03-18
中华养生保健(2020年4期)2020-11-16
山东医药(2020年9期)2020-05-20
科学技术创新(2020年4期)2020-01-08
