
miR-30a靶向Beclin1蛋白在结肠癌中的作用及其机制*
2020-08-23 10:15刘林孟涛雷程杨新辉王海江
贵州医科大学学报 2020年8期
刘林,孟涛,雷程,杨新辉,王海江
(新疆医科大学附属肿瘤医院 胃肠外科,新疆 乌鲁木齐 830011)
结直肠癌(colorectal cancer,CRC)是严重威胁人类健康的恶性肿瘤,据统计每年全世界CRC发病人数约120万,大约有60万人因此丧命[1-2],我国CRC人群的死亡率在十年间已经增加了63.4%[3],而导致CRC患者死亡的主要原因是癌细胞发生转移,但其具体的调控机制尚不十分清楚[4-5]。已有研究表明,有多种功能性miRNA在CRC病理进程中发挥调节作用。miRNA是一类单链内源性非编码小分子RNA,通过与靶基因的3′UTR特异性结合,使mRNA降解或抑制蛋白质翻译过程,进而调控靶基因的蛋白表达水平[6]。目前已发现多种miRNAs可以参与到肿瘤细胞增殖、侵袭、凋亡和自噬等生物学行为[7-8]。细胞自噬(autophagy)是生物体内存在的一种高度保守的代谢过程,主要通过吞噬细胞内受损、衰老或失去功能的蛋白质、或细胞器形成自噬泡,自噬泡可被溶酶体中的酶水解为细胞生存所必需的成分[9-10]。细胞自噬与肿瘤的生成和癌症的发展有关,有实验表明抑制细胞自噬可以恢复治疗的敏感性,增加癌细胞的死亡[11-12]。机体的自噬过程需要多个自噬调节蛋白的参与,其中Beclin1可以介导其它自噬相关蛋白定位于自噬泡,在肿瘤的发生、发展、预后及治疗等方面发挥重要功能[13]。目前,有少数关于Beclin1在结肠癌细胞的生长和转移中的相关研究,但关于结肠癌、miRNA和自噬之间相互作用的研究较少,因此本课题以结肠癌细胞为研究对象,采用MTT、流式细胞术和侵袭迁移实验检测miR-30a对结肠癌细胞增殖、凋亡、侵袭能力以及化疗敏感性的影响,并采用双荧光素酶实验和Western blot等实验验证miR-30a对Beclin1的调控作用,为深入研究结肠癌中miR-30a的调控机制提供依据。
1 材料与方法
1.1主要材料与试剂
选取人结肠癌细胞株HCT116由新疆医科大学附属肿瘤医院实验室提供,miR-30a慢病毒表达载体GV369委托上海吉凯基因公司设计合成,Luc荧光素酶质粒购自上海吉凯基因化学技术有限公司(编号GV306),转染试剂由吉凯公司内部配制。U6、miR-30a、Beclin1-WT及Beclin1-Mut引物由上海生工有限公司合成,DMEM(高糖)培养基、胎牛血清、MEM培养基、Lipo 2000阳离子脂质体购自Invitrogen公司,兔抗Beclin1和兔抗GAPDH抗体购自Abgent公司,BCA蛋白定量试剂盒、RIPA裂解液、HRP标记羊抗兔二抗购自南京碧云天生物公司,Trizol购自TaKaRa公司,质粒提取试剂盒购自天根生化科技有限公司,SYBR Green PCR Kit购自Qiagen公司,基底膜基质凝胶Matrigel购自美国BD公司。
1.2方法
1.2.1细胞培养 人结肠癌细胞系HCT116使用含有10%小牛血清D的MEN培养液在37 ℃含有5% CO2的细胞培养箱内培养,按照ATCC说明书建议方式操作,取指数生长期的细胞用于实验。
1.2.2慢病毒的获得 使用三载体系统生产病毒,转染前先培养293T细胞至指数生长期,用胰蛋白酶消化后重悬至细胞密度为1×109/L,重新接种于6孔细胞培养皿置于细胞培养箱备用;按照说明书配置质粒混合液(20 μg GV369-miR-30a或GV369-NC、15 μg pHelper 1.0和10 μg pHelper 2.0);加入5% Lipofectamine 2000转染试剂3 mL到293T细胞,常规培养12 h;弃去培养基,用4 ℃预冷的PBS洗2次,更换DMEM完全培养基,置于细胞培养箱;24 h后收集细胞上清,经微孔滤膜过滤及高速离心后得到病毒浓缩液。
1.2.3细胞感染及分组 参考文献[14],HCT-116细胞经胰蛋白酶消化后接种到细胞培养6孔板,每孔105个细胞;次日细胞增殖至20%~30%密度进行转染,用100 mg/L的polybrene稀释病毒,加入到HCT-116细胞进行感染,常规培养12 h后,用完全培养基更换掉原培养基;常规培养3 d,加入嘌呤霉素(终浓度为2 mg/L)进行筛选,对细胞计数,统计感染效率。将筛选得到的能够稳定转染GV369-miR-30a和GV369-NC的HCT-116细胞分别命名为miR-30a和NC。实验设置空白对照组CON(未经病毒感染的HCT-116细胞)、阴性细胞组NC(转染GV369-NC的HCT-116细胞)、阳性细胞组miR-30a(转染GV369-miR-30a的HCT-116细胞),每组分别用5 μmol/L奥沙利铂注射液处理或不处理。
1.2.4细胞中目的基因检测 HCT-116细胞在消化后,用Trizol提取细胞内的总RNA,并对RNA的纯度及浓度进行测定,使用逆转录试剂盒进行cDNA的合成,反应程序:37 ℃,12 min;84 ℃,6 s。应用实时荧光定量PCR(qRT-PCR)方法检测各组细胞中目的基因的表达水平,采用2-ΔΔCt法、U6为内参基因进行相对定量,每组实验重复3次,引物序列见表1。

表1 qRT-PCR引物序列Tab.1 Primer sequences of qRT-PCR
1.2.5细胞增殖实验 将空白对照组CON、NC组和miR-30a组细胞分别制备成悬液(密度为2×107/L),分别取100 μL加入到96孔板的每个孔里,每孔加入5 g/L的MTT溶液10 μL,孵育4 h;细胞变成紫黑色后吸取培养基,PBS清洗3次后置于显微镜下观察,记数。
1.2.6Transwell迁移实验 用BD公司的Matrigel 1 ∶8稀释制备凝胶,HCT-116细胞经胰蛋白酶消化后,用MEM培养基清洗3次,再用含BSA的无血清培养基重悬至5×108/L;每个小室加100 μL细胞悬液,取600 μL培养基(含有20%FBS)加入到24孔板下室,37 ℃培养24 h;弃去培养液,用PBS(经37 ℃预热)清洗3次,再加入4%多聚甲醛(经37 ℃预热)固定30 min;PBS清洗3次,将小室晾干;每孔加入0.1%结晶紫染色20 min,用棉签擦去小室内未迁移的细胞,置于显微镜下观察并记数。
1.2.7流式细胞术 参考文献[14],将处于指数生长期的HCT-116细胞用胰蛋白酶进行消化,用MEM完全培养液调节细胞悬液浓度为109/L,加入预冷的75%乙醇进行细胞固定,震荡混匀后室温静置10 min;去除75%乙醇,用预冷的PBS溶液清洗3次,震荡混匀后室温静置10 min;将HCT-116细胞重悬于binding buffer,加入Annexin-V-FITC(40 mg/L)1 μL,混匀后避光在冰上静置15 min;加入RNA酶轻轻震荡混匀,室温静置5 min后,将HCT-116细胞转移到流式检测管中,向其中加入的PBS 500 μL,上机前加入PI(100 mg/L)0.5 μL,注意避光操作。
1.2.8靶基因预测、载体构建与鉴定 参考文献[14],从miRBase数据库查找miR-30a序列,获取其前体和成熟体的RNA序列;使用PicTar、TargetScan、miRanda等生物信息学软件对miR-30a的靶基因进行预测,结合文献获得的信息,最终选定Beclin1开展研究。miR-30a能够与Beclin1的基因编码3′UTR区的位点特异性结合,为了进一步验证上述的预测结果,委托生物公司构建了含有野生型Beclin1(Beclin1-WT)和3′UTR区突变的Beclin1(Beclin1-Mut)的荧光素酶报告基因载体。
1.2.9双荧光素酶报告基因实验 通过Lipo2000将psi-CHECK-Beclin1-WT、psi-CHECK-Beclin1-Mut分别和GV369-miR-30a或GV369-NC共转染到HCT-116细胞中,置于培养箱6 h后换完全培养基;48 h后,去除培养基,用PBS洗涤,加入RIPA裂解液后用枪头吹打,低温离心后转移上清;使用DLRTM系统检测各组细胞中萤火虫(photinus pyralis)荧光素酶和海肾(renilla reniformis)荧光素酶活性,最终以上述2种荧光素酶的荧光强度的比值来代表细胞的荧光强度。
1.2.10Western blot 各组细胞去除培养液后加入RIPA细胞裂解液(含有PMSF 1 μL和蛋白酶抑制剂 1 μL)裂解,分别提取总蛋白;根据BCA法测定蛋白浓度,沸水浴煮5 min后离心取上清上样;电泳分离后,将蛋白电转至PVDF膜,用封闭液(5%奶粉)室温封闭2 h;加入一抗anti-Beclin(1 ∶2 000)或anti-GAPDH(1 ∶2 000),4℃振荡过夜;TBST洗10 min(3次),加入HRP标记的二抗,室温振荡1 h,TBST洗3次后进行化学发光显影。
1.3统计学分析
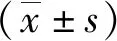
2 结果
2.1慢病毒感染结肠癌细胞的效率
观察miR-30a组和NC组细胞,可见约80%以上的HCT-116细胞呈现绿色荧光,表明miR-30a慢病毒载体感染成功(图1A)。qRT-PCR结果显示,miR-30a组细胞的miR-30a表达水平明显高于NC组,表达丰度是NC组的1 408倍,差异有统计学意义(P<0.001,图1B)。
注:A为慢病毒感染的HCT-116细胞的效率检测;B为慢病毒感染HCT-116细胞后miR-30a的表达水平;(1)与NC组比较,P<0.001。图1 转染后HCT116细胞中miR-30a的表达水平Fig.1 Expression level of miR-30a in HCT116 cells after transfecting
2.2miR-30a过表达抑制结肠癌细胞的增殖能力
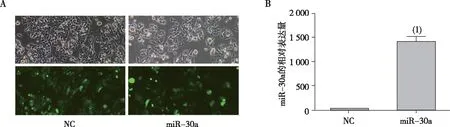
MTT细胞增殖抑制实验结果表明,与空白对照组和NC组相比,miR-30a组的HCT-116细胞增殖能力受到显著抑制,差异有统计学意义(P<0.05);在加入奥沙利铂注射液后,各组细胞的增殖能力更加显著被抑制,而与对照组相比,miR-30a过表达的细胞对奥沙利铂注射液的敏感性增强,差异有统计学意义(P<0.05)。见图2。
注:1、2、3、4、5及6分别为空白对照组、NC组、miR-30a组、对空白对照组用奥沙利铂处理、NC组用奥沙利铂处理及miR-30a组奥沙利铂处理;(1) 与NC组相比,P<0.05。图2 miR-30a过表达对HCT-116细胞增殖能力的影响Fig.2 Effect of miR-30a over-expression on the proliferation of HCT116 cells
2.3miR-30a过表达抑制结肠癌细胞的迁移能力
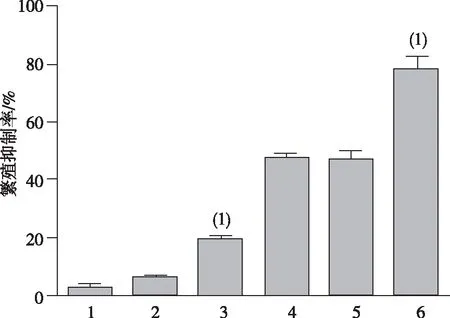
细胞迁移实验结果表明,无论是否用奥沙利铂注射液处理,miR-30a组HCT-116细胞在血清的诱导下迁移通过聚碳酸酯膜的数量显著低于空白对照组和NC组细胞,差异有统计学意义(P<0.05);而在加入奥沙利铂注射液后,3组细胞的迁移能力进一步下降,而miR-30a组细胞的迁移能力仍低于两个对照组,差异有统计学意义(P<0.05)。见图3。
2.4miR-30a过表达对HCT116细胞凋亡的作用
流式细胞术的检测结果显示,空白对照组HCT116细胞的凋亡比例为(1.80±0.17)%,阴性对照组为(1.82±0.05)%,二者间差异无统计学意义(P>0.05);miR-30a组细胞凋亡比例为(3.86±0.09)%,与2个对照组相比,差异有统计学意义(P<0.01)。加入奥沙利铂注射液后,miR-30a组细胞凋亡比例显著增加至(18.24±0.56)%,显著高于对照组细胞,差异有统计学意义(P<0.01),见图4。
2.5双荧光素酶报告系统
如图5所示,与对照组相比(Beclin1-WT + NC),Beclin1-WT和miR-30a共表达导致荧光素酶的活性显著下降,荧光活性下降(28.2%±4.1)%,差异有统计学意义(P<0.05);而当Beclin1-Mut与miR-30a共表达并不能显著影响荧光素酶的活性,与Beclin1-Mut + miR-30a相比,差异无统计学意义(P>0.05)。
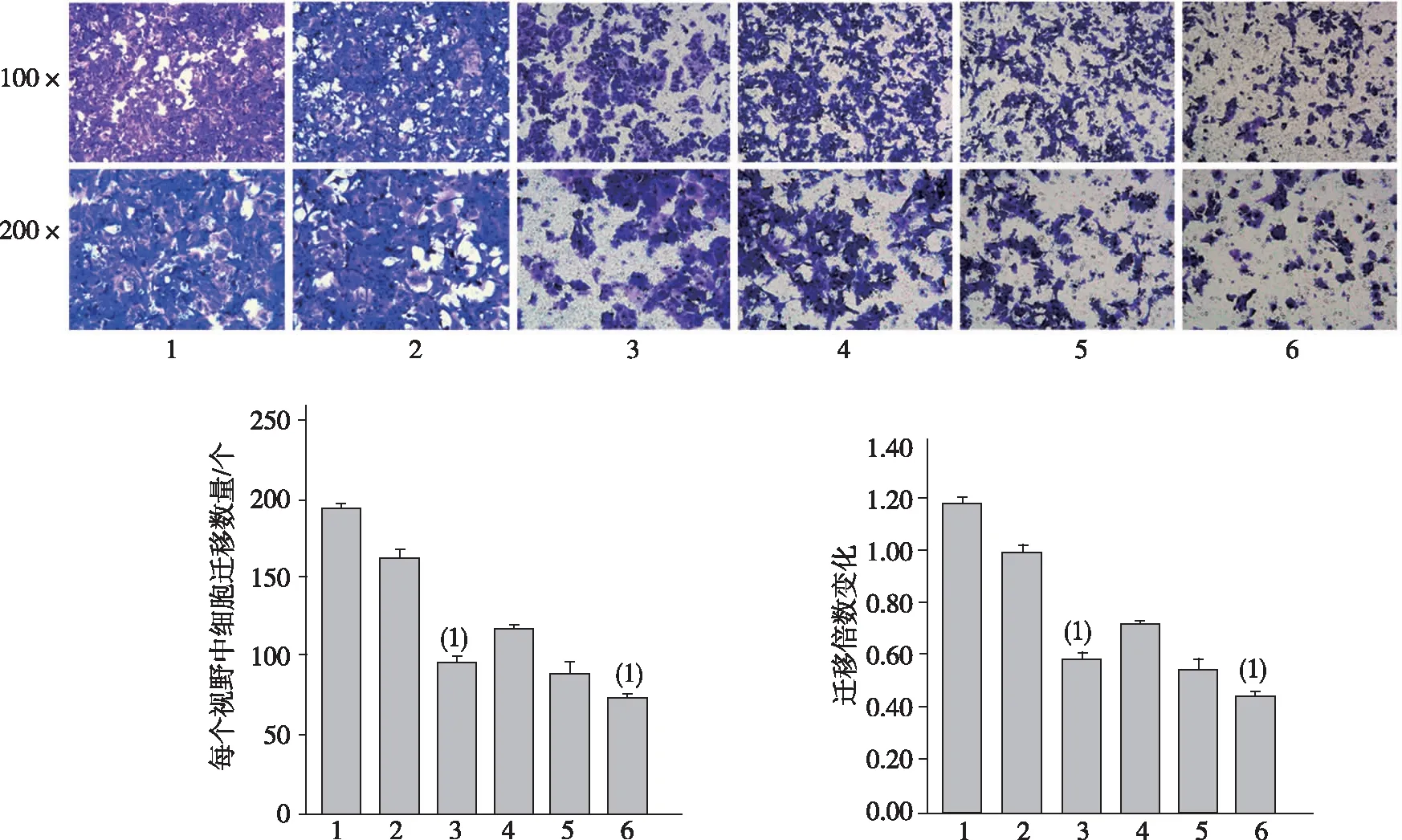
注:1、2、3、4、5及6分别为空白对照组、NC组、miR-30a组、对空白对照组用奥沙利铂处理、NC组用奥沙利铂处理及miR-30a组奥沙利铂处理;(1) 与NC组相比,P<0.05。 图3 miR-30a过表达对HCT-116细胞迁移能力的影响Fig.3 Effect of miR-30a over-expression on migration of HCT116 cells
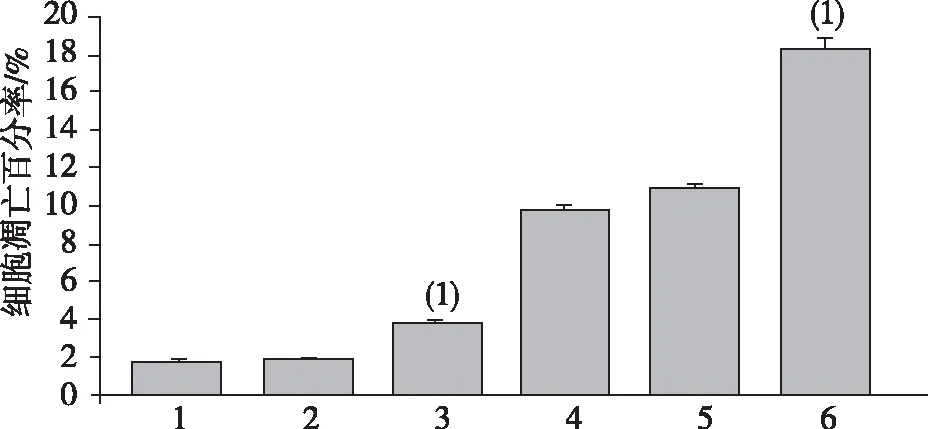
注:1、2、3、4、5及6分别为空白对照组、NC组、miR-30a组、对空白对照组用奥沙利铂处理、NC组用奥沙利铂处理及miR-30a组奥沙利铂处理;与NC组相比,(1) P<0.01。图4 miR-30a表达对HCT116细胞凋亡的作用Fig.4 Effect of miR-30a expression on apoptosis of HCT116 cells
2.6miR-30a过表达对Beclin1 基因mRNA和蛋白表达的影响
qRT-PCR结果显示,与空白对照组和NC相比,miR-30a组的Beclin1表达水平显著下调,差异有统计学意义(P<0.01)。在加入奥沙利铂注射液后,各组细胞的Beclin1表达均出现增加,奥沙利铂处理的NC组中Beclin1的表达量相较于处理前上升了(5.67±0.9)倍;奥沙利铂处理的miR-30a组与NC组相比,Beclin1表达量下降了(34.5±6.8)%,差异有统计学意义(P<0.01),见图6A。如图6B所示,Western blot结果与RT-qPCR检测结果一致:无论奥沙利铂处理与否,与空白对照组和NC组相比,miR-30a组细胞中Beclin1蛋白的表达均下降;在加入奥沙利铂注射液后,各组细胞中Beclin1蛋白表达量均出现增加。
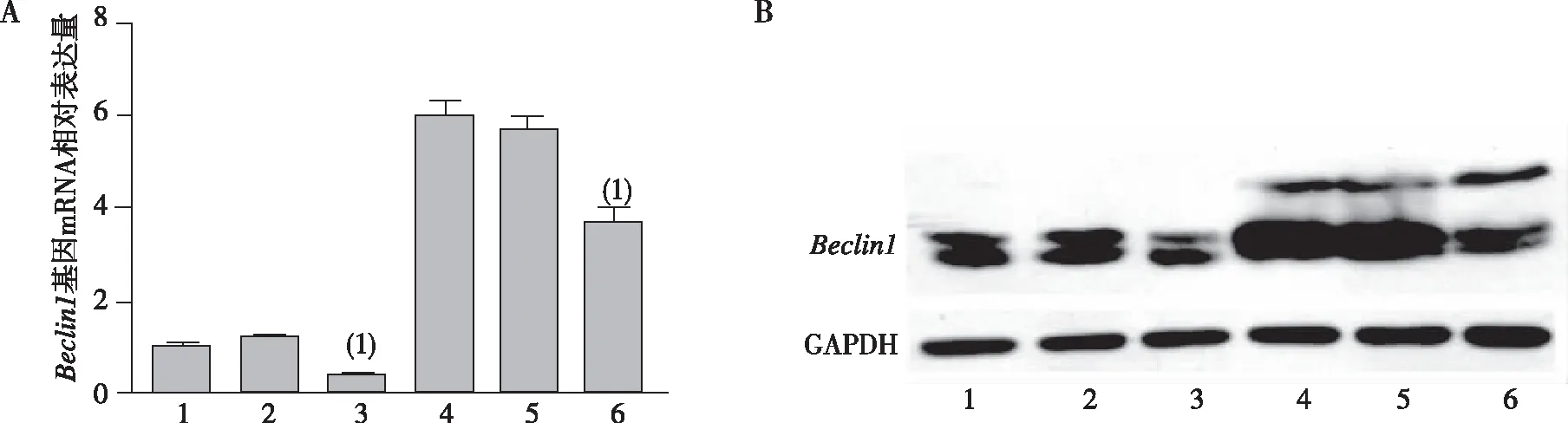
注:A为Beclin1基因mRNA的相对表达水平,B为Beclin1蛋白的表达水平;1、2、3、4、5及6分别为空白对照组、NC组、miR-30a组、对空白对照组用奥沙利铂处理、NC组用奥沙利铂处理及miR-30a组奥沙利铂处理;(1) 与NC组相比,P<0.01。图6 Beclin1的mRNA和蛋白的表达水平Fig.6 Expression level of mRNA and albumen on Beclin1
3 讨论
研究表明,miRNAs能够调控多种癌细胞中特定基因的表达,也可直接作为癌基因或抑癌基因直接参与到肿瘤的发生发展过程,因此,miRNAs被认为是肿瘤治疗的潜在靶分子[14-15]。有研究发现,转染miR-612的膀胱癌细胞生长、增殖、侵袭能力会受到抑制,也会抑制EMT进程,进而抑制癌细胞的侵袭和转移[16];同样发现miR-221/222也可以抑制肿瘤细胞EMT进程[17]。LncRNA NEAT1可以通过靶向miR-34a促进自噬,进而促进结肠癌细胞对5-氟尿嘧啶的抗性,而过表达miR-34a可以提高细胞的敏感性[18]。lncRNA-ROR可以通过抑制p53信号通路进而促进鼻咽癌细胞的增殖,迁移和药物抗性[19];也有研究表明miR-630可以抑制乳腺癌的发展[20]。
Beclin1,属于酵母ATG6的同系物,是最早被鉴定为具有调控自噬的发生的蛋白,在恶性肿瘤中发挥了重要作用,被认为是具有抑癌功能的基因,可以诱导癌细胞发生自噬性死亡[21]。有研究表明miRNA可以通过靶向下调Beclin1的表达来增强癌细胞对化疗的敏感性[22],但是也有研究发现miRNA靶向调控Beclin1的过程与细胞增殖有关[23]。而本研究结果显示,miR-30a的过表达可以下调Beclin1的表达水平,而奥沙利铂则可以促进Beclin1蛋白的表达,并都可以同时抑制结肠癌细胞的增殖、侵袭及凋亡[14],提示miR-30a/Beclin1有可能成为结肠癌治疗的作用靶点。
综上所述,本实验表明在结肠癌细胞中过表达miR-30a,可以通过靶向下调Beclin1的表达来抑制癌细胞增殖,促进细胞凋亡,说明miR-30a有望成为结肠癌靶向治疗的基因分子。
猜你喜欢
中日友好医院学报(2022年4期)2022-10-15
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
看世界·学术下半月(2020年7期)2020-09-10
山东医药(2020年9期)2020-05-20
广东农业科学(2017年10期)2018-01-25
中国医学创新(2017年22期)2017-11-15
中国组织化学与细胞化学杂志(2016年4期)2016-02-27
西南军医(2016年2期)2016-01-23
中国医药导报(2015年27期)2015-02-28
