
miRNA-34a 靶向调控叉头框蛋白P1对活化B细胞样弥漫性大B细胞淋巴瘤细胞凋亡的影响
2020-07-28 07:30武君梁艳张森森王娟尚念梅杜忠海
癌症进展 2020年12期
武君,梁艳,张森森,王娟,尚念梅,杜忠海
潍坊市中医院肿瘤中心肿瘤科,山东 潍坊 261041
弥漫性大B细胞淋巴瘤(diffuse large B-cell lymphoma,DLBCL)是一种具有高度异质性和高度侵袭性的非霍奇金淋巴瘤(non-Hodgkin lymphoma,NHL),是发病率最高的NHL亚型,约占30%~40%,而在亚洲国家所占比例高达50%[1-2]。Alizadeh等[3]根据基因表达谱(gene expression profiling,GEP)技术将DLBCL分为生发中心B细胞样(germinal center B cell-like,GCB)和活化B细胞样(activated B cell-like,ABC)亚型。研究显示,ABCDLBCL患者的预后比GCB-DLBCL差,仍有30%~40%的ABC-DLBCL患者接受现有的方案治疗,无法得到持续缓解[4-5]。因此,研究DLBCL发生发展的生物学异质性,寻找新的生物治疗靶点对DLBCL的临床诊断和治疗有积极意义。近年来研究发现,微小RNA(microRNA,miRNA)在人类肿瘤、胚胎发育及代谢疾病等多种疾病中都发挥重要作用[6-7]。其中miRNA-34a作为抑癌基因p53的下游基因,与多种恶性肿瘤的发生发展密切相关[8-11]。miRNA-34a转录受p53蛋白的正调控,引起细胞周期变化或细胞凋亡等,在p53作用通路中发挥重要作用[12-14]。研究表明,miRNA-34a在包括DLBCL在内的多种恶性肿瘤中低表达[15],提示miRNA-34a可能是潜在的抑癌因子。已有研究表明,miRNA-34a可以促进DLBCL细胞凋亡从而抑制DLBCL的发展[16]。叉头框蛋白 P1(forkhead box protein 1,FOXP1)是FOXP亚家族转录因子成员,广泛表达于各个组织并发挥多种生理功能,在不同的肿瘤组织中发挥不同的生物学作用[17]。FOXP1是B细胞发育过程中的关键转录调节因子,表达异常时容易产生B细胞淋巴瘤[18]。FOXP1蛋白在ABC-DLBCL中高表达,提示FOXP1可能作为癌基因参与了DLBCL的发生发展[19]。Cerna等[20]研究发现,在慢性淋巴细胞性白血病中,FOXP1可能作为miRNA-34a的靶点,通过负调控FOXP1蛋白的表达来限制B细胞受体信号通路。但miRNA-34a引起DLBCL细胞凋亡的具体机制目前尚不明确。因此,本研究探讨miRNA-34a和FOXP1的表达变化对ABC-DLBCL细胞增殖和凋亡的影响,探讨相关分子作用机制,现报道如下。
1 材料及方法
1.1 细胞、主要试剂和仪器
人胚肾细胞株293T、ABC-DLBCL细胞株SUDHL-2均购自上海泽叶生物科技有限公司。胎牛血清、RPMI-1640培养基、流式检测试剂盒均购自美国Thermo Fisher公司,脂质体LipofectamineTM2000、Trizol试剂均购自美国Invitrogen公司,FOXP13'非翻译区(3'-untranslated region,3'-UTR)野生型、突变型报告基因载体及其空载体均购自上海吉满生物科技有限公司,miRNA-34a mimics购自美国Invitrogen公司,FOXP1siRNA购自美国Sigma-Aldrich公司,逆转录试剂、实时荧光定量逆转录聚合酶链反应(reverse transcription-polymerase chain reaction,RT-PCR)试剂盒均购自德国Qiagen公司,Bradford蛋白浓度检测试剂盒购自上海碧云天生物技术有限公司,抗FOXP1抗体、抗甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗体、辣根过氧化物酶(horseradish peroxidase,HRP)偶联二抗均购自美国CST公司,聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜购自美国Pharrnacai公司,双荧光素酶报告基因试剂盒购自美国Promega公司,荧光定量PCR检测引物由上海生工生物工程有限公司合成。
1.2 实验方法
1.2.1 细胞培养、转染及分组 人胚肾细胞株293T、ABC-DLBCL细胞株SU-DHL-2分别在5%CO2、37℃条件下,于含10%胎牛血清的RPMI-1640培养基中培养。将细胞以1×105/孔接种于24孔板,当细胞的生长密度为60%左右时开始进行转染。按照LipofectamineTM2000试剂说明书中步骤及试剂使用量进行常规转染:将6 μl脂质体及20 pmol的复合物分别稀释于250 μl的无血清培养基中,室温静置5 min后轻柔混匀,室温静置15 min后分别均匀加入各组培养板中,使复合物的终浓度为10 nmol/L,37℃温箱中培养6 h后换为完全培养基常规培养。根据SU-DHL-2细胞不同处理因素进行分组,其中未经转染处理的细胞作为Blank组,转染miRNA-34a mimics的细胞作为miRNA-34a过表达组,转染FOXP1siRNA的细胞作为FOXP1基因沉默组,转染一段无义序列的细胞作为阴性对照NC组,转染后常规培养48 h。
1.2.2 双荧光素酶报告基因实验 通过Targetscan、miRanda和miRDB生物信息数据库分析和既往相关研究报道提示FOXP1的3'-UTR区是miRNA-34a结合靶位点之一。因此,本研究通过双荧光素酶报告基因实验验证miRNA-34a对FOXP1野生型3'-UTR特异性结合的作用,将人胚肾细胞株293T根据实验设计进行转染分组,组1,转染FOXP13'-UTR无义序列+miRNA无义序列;组2,转染FOXP13'-UTR无义序列+miRNA-34a mimics;组3,转染FOXP13'-UTR野生报告质粒+miRNA无义序列;组4,转染FOXP13'-UTR野生报告质粒+miRNA-34a mimics;组5,转染FOXP13'-UTR突变报告质粒+miRNA无义序列;组6,转染FOXP13'-UTR突变报告质粒+miRNA-34a mimics。转染后分别常规培养24 h,然后按照双荧光素酶报告基因检测试剂盒(Promega)说明书步骤处理细胞,在多功能微孔板检测仪上测定并分析结果。
1.2.3 RT-PCR检测miRNA-34a和FOXP1mRNA的相对表达量 分别取Blank组、NC组、50 nmol/L miRNA-34a过表达组、100 nmol/L miRNA-34a过表达组、50 nmol/LFOXP1基因沉默组、100 nmol/LFOXP1基因沉默组SU-DHL-2细胞。采用Trizol法分别提取各组细胞总RNA,并检测其浓度及纯度;按照逆转录试剂盒说明书反转录成cDNA,并以此为模板于ABI PRISM 7900HT型荧光定量PCR仪上行实时PCR反应,以U6和GAPDH为内参。PCR反应体系4 μl,反应条件:95 ℃ 5 min,95 ℃10 s,60 ℃ 20 s,50个循环;95 ℃ 10 s,60 ℃ 10 s,40 ℃ 30 s。采用2-ΔΔCt法检测各组细胞miRNA-34a和FOXP1mRNA的相对表达量,实验重复3次。引物序列见表1。

表1 引物序列
1.2.4 蛋白质印迹法(Western blot)检测FOXP1蛋白的相对表达量 分别取Blank组、NC组、50 nmol/L miRNA-34a过表达组、100 nmol/L miRNA-34a过表达组、50 nmol/LFOXP1基因沉默组、100 nmol/LFOXP1基因沉默组SU-DHL-2细胞。加入适量的放射免疫沉淀(radio-immunoprecipitation assay,RIPA)裂解液,提取总蛋白,Bradford法检测蛋白浓度。取50 ng蛋白置于离心管中加入适量十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离蛋白,转移至PVDF膜,TBST洗膜3次,5%脱脂牛奶封闭2 h,加入一抗(稀释浓度为1∶1000)4℃过夜;TBST洗涤3次,加入二抗,室温孵育2 h;化学发光显色法显影,以GAPDH为内参,检测各组细胞FOXP1蛋白的相对表达量,实验重复3次。
1.2.5 CCK8法检测细胞增殖能力 选取Blank组、NC组及相对表达量较高的100 nmol/L miRNA-34a过表达组和FOXP1基因沉默组SU-DHL-2细胞进行检测。以3×103/孔接种于96孔板,分别在0、24、48、72、96 h时加入10 μl的CCK8溶液,每组设置6个重复孔;37℃培养2 h后,使用多功能微孔板检测仪检测450 nm处各孔吸光度(optical density,OD)值,记录结果并绘制细胞生长曲线。实验重复3次。
1.2.6 流式细胞仪检测细胞凋亡情况 同增殖实验选取Blank组、NC组及相对表达量较高的100 nmol/L miRNA-34a过表达组和FOXP1基因沉默组SU-DHL-2细胞进行检测。采用预冷磷酸盐缓冲液洗涤2次,加入结合缓冲液制成细胞悬液;按试剂盒说明书中建议的试剂体系用量依次加入膜联蛋白V(AnnexinⅤ)、碘化丙啶(propidium iodide,PI)后轻轻混匀,室温避光处孵育15 min,流式细胞仪检测细胞凋亡情况,Cell Quest软件进行结果分析。实验重复3次。
1.3 统计学方法
采用SPSS 21.0软件对所有数据进行统计分析,计量资料以均数±标准差(±s)表示,多组间比较采用单因素方差分析,两组间比较采用t检验;以P<0.05为差异有统计学意义。
2 结果
2.1 miRNA-34a的相对表达量的比较
NC组和Blank组细胞中miRNA-34a相对表达量比较,差异无统计学意义(P>0.05)。50 nmol/L miRNA-34a过表达组和100 nmol/L miRNA-34a过表达组SU-DHL-2细胞中miRNA-34a的相对表达量均明显高于Blank组和NC组细胞,其中100 nmol/L miRNA-34a过表达组SU-DHL-2细胞中miRNA-34a的相对表达量明显高于50 nmol/L miRNA-34a过表达组细胞,差异均有统计学意义(P<0.01)。(表2)
表2 各组SU-DHL-2细胞miRNA-34a相对表达量的比较(±s)

表2 各组SU-DHL-2细胞miRNA-34a相对表达量的比较(±s)
注:a与Blank组比较,P<0.01;b与NC组比较,P<0.01;c与50 nmol/L miRNA-34a过表达组比较,P<0.01
组别Blank组NC组50 nmol/L miRNA-34a过表达组100 nmol/L miRNA-34a过表达组1.00±0.09 1.13±0.10 4.81±0.15a b 10.67±0.17a b c miRNA-34a相对表达量
2.2 miRNA-34a过表达对SU-DHL-2细胞增殖和凋亡的影响
研究发现随着转染时间的延长,miRNA-34a过表达组、NC组和Blank组SU-DHL-2细胞OD均逐渐升高,但转染同一时间点3组SU-DHL-2细胞OD值比较,差异均无统计学意义(P>0.05)(表3)。miRNA-34a过表达组SU-DHL-2细胞凋亡率(32.76±0.19)%,明显高于NC组细胞的(20.87±0.14)%,差异有统计学意义(t=81.597,P=0.000)(图1)。
表3 不同转染时间各组SU-DHL-2细胞的OD值(±s)

表3 不同转染时间各组SU-DHL-2细胞的OD值(±s)
转染时间0 h 24 h 48 h 72 h 96 h 0.185±0.03 0.384±0.06 0.585±0.06 0.705±0.06 0.868±0.07 0.195±0.04 0.379±0.05 0.576±0.05 0.695±0.06 0.858±0.07 0.187±0.02 0.403±0.07 0.689±0.08 0.787±0.08 0.997±0.09 Blank组NC组miRNA-34a过表达组

图1 流式细胞仪检测miRNA-34a过表达对SU-DHL-2细胞凋亡的影响
2.3 miRNA-34a过表达对FOXP1mRNA和FOXP1蛋白表达的影响
Blank组和NC组SU-DHL-2细胞中FOXP1mRNA和FOXP1蛋白相对表达量比较,差异均无统计学意义(P>0.05)。miRNA-34a过表达组细胞FOXP1mRNA和FOXP1蛋白的相对表达量均明显低于Blank组和NC组细胞,差异均有统计学意义(P<0.01)。(表4)
表4 miRNA-34a过表达对FOXP1 mRNA和FOXP1蛋白表达的影响(±s)

表4 miRNA-34a过表达对FOXP1 mRNA和FOXP1蛋白表达的影响(±s)
注:a与Blank组比较,P<0.01;b与NC组比较,P<0.01
组别Blank组NC组miRNA-34a过表达组1.00±0.08 0.98±0.08 0.25±0.06a b 1.02±0.10 0.94±0.09 0.18±0.04a b FOXP1 mRNA FOXP1蛋白
2.4 双荧光素酶报告基因检测结果
双荧光素酶报告基因实验验证miRNA-34a对FOXP1野生型3'-UTR特异性结合的作用,结果显示,组1的荧光素酶活性值为(1.00±0.03)与组2的(0.98±0.02)比较,差异无统计学差异(t=0.961,P=0.196);组3的荧光素酶活性值为(0.95±0.09),高于组 4的(0.28±0.02),差异有统计学意义(t=12.587,P=0.000);组5的荧光素酶活性值为(0.93±0.04)与组6的(0.99±0.04)比较,差异无统计学意义(t=1.837,P=0.070)。(图2)
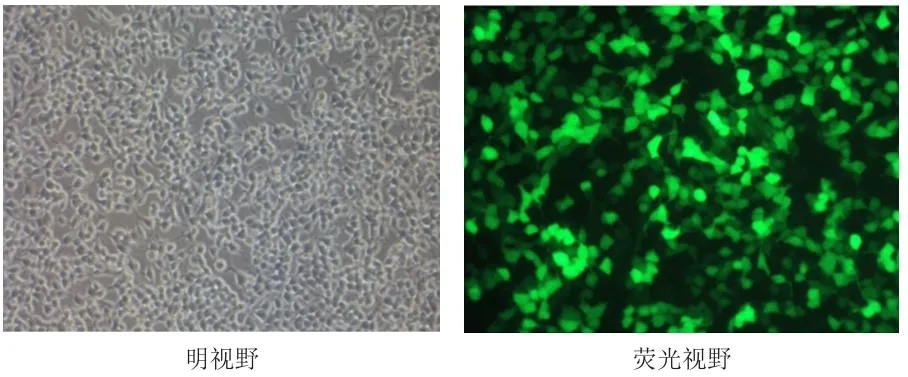
图2 FOXP13'-UTR 野生型荧光素酶质粒转染293T细胞图(×200)
2.5 FOXP1siRNA对FOXP1mRNA和FOXP1蛋白表达水平的影响
Blank组与NC组细胞FOXP1mRNA及FOXP1蛋白的相对表达量比较,差异均无统计学意义(P>0.05)。50 nmol/LFOXP1基因沉默组和100 nmol/LFOXP1基因沉默组细胞FOXP1mRNA及FOXP1蛋白的相对表达量均明显低于Blank组和NC组细胞,其中100 nmol/LFOXP1基因沉默组FOXP1mRNA及FOXP1蛋白的相对表达量,均明显低于50 nmol/LFOXP1基因沉默组细胞,差异均有统计学意义(P<0.01)。(表5)
2.6 FOXP1基因沉默对SU-DHL-2细胞增殖和凋亡的影响
随着转染时间的延长,Blank组、NC组及FOXP1基因沉默组SU-DHL-2细胞OD值均逐渐升高,但相同转染时间点3组细胞OD值比较,差异无统计学意义(P>0.05)(表6);FOXP1基因沉默组SU-DHL-2细胞凋亡率(35.18±0.22)%,明显高于NC组细胞的(21.98±0.16)%,差异有统计学意义(t=84.046,P=0.000)(图3)。
表5 FOXP1 siRNA对FOXP1 mRNA和FOXP1蛋白表达水平的影响(±s)

表5 FOXP1 siRNA对FOXP1 mRNA和FOXP1蛋白表达水平的影响(±s)
注:a与Blank组比较,P<0.01;b与NC组比较,P<0.01;c与50 nmol/L FOXP1基因沉默组比较,P<0.01
组别Blank组NC组50 nmol/L FOXP1基因沉默组100 nmol/L FOXP1基因沉默组1.00±0.03 1.01±0.05 0.40±0.03a b 0.09±0.01a b c 0.98±0.03 0.87±0.05 0.31±0.06a b 0.13±0.04a b c FOXP1 mRNA FOXP1蛋白
表6 CCK8法检测转染FOXP1 siRNA后SU-DHL-2细胞增殖情况(±s)

表6 CCK8法检测转染FOXP1 siRNA后SU-DHL-2细胞增殖情况(±s)
Blank组0.185±0.03 0.395±0.04 0.605±0.03 0.753±0.03 0.918±0.02 0 h 24 h 48 h 72 h 96 h 0.195±0.03 0.390±0.03 0.598±0.02 0.747±0.04 0.911±0.04 0.184±0.04 0.377±0.02 0.567±0.08 0.739±0.05 0.898±0.02转染时间NC组FOXP1基因沉默组

图3 流式细胞仪检测FOXP1siRNA对SU-DHL-2细胞凋亡的影响
3 讨论
研究表明,细胞发生了非正常的增殖和凋亡是导致肿瘤发生发展的重要原因之一[21]。由于DLBCL是一类在分子病理学、细胞遗传学、临床表现及预后等多个方面都具有高度异质性的恶性肿瘤,多种因素、多种基因交互参与其中,因此,DLBCL发生发展过程中所涉及的分子机制仍不完全明确,有待于进行更深入的研究[22]。因此,进一步探索与DLBCL发生发展、临床疗效或预后相关的新的分子标志物、开发特异性治疗靶点、优化临床治疗策略,对患者进行预后分层及个体化治疗,对改善临床疗效,提高患者的生存率有重要意义。
既往研究表明,miRNA常通过与染色体脆性位点或与肿瘤相关的基因组区结合,起到调节靶基因转录后表达水平的作用[23]。Yu和Li[24]研究发现,淋巴瘤患者存在明显的miRNA表达失调,表明miRNA对淋巴瘤的发生发展可能有重要作用。多项研究结果表明,miRNA-17-92、miRNA-21、miRNA-155等在DLBCL中高表达,直接参与了DLBCL的发生和发展过程[25-27]。miRNA可能作为分子标志物及个体化治疗的潜在靶点,将对DLBCL的治疗和预后评估产生重要影响。经典抑癌基因p53的直接下游靶标miRNA-34a在ABC-DLBCL细胞中相对表达量很低,人为上调miRNA-34a的表达后可以诱导细胞凋亡和细胞周期停滞,但具体的分子机制仍不明确[28-29]。本研究将miRNA-34a mimics转染至SU-DHL-2细胞,观察miRNA-34a过表达对SU-DHL-2细胞增殖和凋亡的影响,结果显示,miRNA-34a过表达组细胞的凋亡率高于NC组,人为上调miRNA-34a的表达后对SU-DHL-2细胞的增殖能力没有影响,与以上结论相符。
本研究结合既往研究结果及Targetscan、mi-Randa和miRDB三个生物学数据库筛选出FOXP1作为进一步研究对象。Gascoyne和Banham[30]研究结果显示,FOXP1基因表达失调在DLBCL的发病机制中可能发挥重要作用。但目前关于FOXP1在DLBCL中作用机制的研究报道较少。本研究通过双荧光素酶报告基因检测结果显示,组3(转染FOXP13'-UTR野生报告质粒+miRNA无义序列)荧光素酶活性值高于组4(转染FOXP1 3'-UTR野生报告质粒+miRNA-34a mimics),差异有统计学意义(P<0.01),表明miRNA-34a过表达与FOXP13'-UTR(野生型)能够特异性结合从而降低荧光素酶活性值,FOXP1是miRNA-34a直接调控的靶基因。通过进一步的RT-PCR和Western blot实验结果显示,与Blank组和NC组比较,miRNA-34a过表达组中FOXP1mRNA及FOXP1蛋白相对表达量均受到明显抑制,表明miRNA-34a对FOXP1起到负调控作用,miRNA-34a过表达明显抑制了SUDHL-2细胞FOXP1转录后的表达水平;这与其他多项研究结果相一致[20,31-33]。在进一步FOXP1基因沉默对SU-DHL-2细胞增殖和凋亡影响的研究中发现,FOXP1基因沉默对SU-DHL-2细胞凋亡有明显的促进作用,认为miRNA-34a可能通过负调控FOXP1的表达来促进SU-DHL-2细胞的凋亡,因此,敲除FOXP1可促进SU-DHL-2细胞的凋亡;而FOXP1基因沉默对SU-DHL-2细胞增殖无明显影响。
综上所述,miRNA-34a抑制ABC-DLBCL细胞的发生发展可能与miRNA-34a通过结合FOXP13'-UTR有效抑制FOXP1蛋白表达进而促进细胞凋亡有关。本研究为DLBCL的基因个体化治疗策略提供新的思路和方法,miRNA-34a或FOXP1相互作用可能成为治疗DLBCL的新靶点。
猜你喜欢
湖南畜牧兽医(2021年6期)2022-01-24
食品安全导刊(2021年21期)2021-08-30
天津医科大学学报(2021年4期)2021-08-21
猪业科学(2021年5期)2021-06-02
中国畜禽种业(2021年4期)2021-05-21
西安交通大学学报(医学版)(2021年3期)2021-05-15
广东蚕业(2021年1期)2021-03-18
临床超声医学杂志(2020年12期)2020-12-20
江苏农业科学(2019年23期)2019-03-03
医学信息(2016年29期)2016-11-28
