
植物乳杆菌XN1904E产胞外多糖发酵条件优化
2020-07-08 05:38雷文平刘成国
乳业科学与技术 2020年3期
陈 绮,雷文平,肖 茜,姚 慧,罗 洁,刘成国,周 辉,*
(1.湖南农业大学食品科学技术学院,湖南 长沙 410128;2.湖南省食品科学与生物技术重点实验室,湖南 长沙410128;3.湖南南山牧业有限公司,湖南 邵阳 422512)
乳酸菌是常用于发酵酸乳、饮料等功能性产品的菌种,使用范围广泛。植物乳杆菌是乳酸菌的一种,最适生长温度为35 ℃,最适生长pH值为6.5左右,属于同型发酵乳酸菌[1]。植物乳杆菌胞外多糖是其在生长代谢过程中分泌到细胞壁外溶解于培养环境中的产物[2-3]。乳酸菌胞外多糖是具有国际公认安全地位的物质,并能改善乳酸菌发酵产品的形态、口感和风味[4]。影响乳酸菌代谢产生胞外多糖的因素有很多种,如pH值、温度、碳源、氮源等[5-6]。
张岩春等[7]通过优化产胞外多糖植物乳杆菌C88发酵条件,将植物乳杆菌C88发酵时间延长到24 h,胞外多糖产量从29.86 mg/L提高到40.96 mg/L。陶静等[8]通过单因素试验与正交试验对乳酸菌产胞外多糖培养基组成和生长条件进行优化,结果表明,乳酸乳球菌乳亚种LL9为胞外多糖合成量最高品系,胞外多糖产量可达到262.628 mg/L。Gancel等[9]研究不同糖作为碳源对嗜热链球菌产胞外多糖的影响,结果发现,乳糖作为碳源时胞外多糖生成量最大,这也说明不同菌株的最佳碳源有所差异。胞外多糖条件优化研究中较常用的碳源是葡萄糖、乳糖、蔗糖和半乳糖。
本研究在已有研究基础上进一步研究植物乳杆菌XN1904E产胞外多糖的影响因素。选取培养温度、培养基初始pH值、碳源和氮源4 个单因素,得到最佳单因素水平,进行正交试验,从而得到植物乳杆菌XN1904E产胞外多糖最佳培养条件。
1 材料与方法
1.1 材料与试剂
植物乳杆菌XN1904E 本实验室分离保存。
M R S 肉汤(含蛋白胨1 0.0 g、牛肉粉5.0 g、酵母粉4.0 g、柠檬酸三胺2.0 g、葡萄糖20.0 g、吐温-80 1.0 mL、乙酸钠5.0 g、磷酸氢二钾2.0 g、硫酸镁0.2 g、硫酸锰0.05 g、蒸馏水1 L,pH 6.4、121 ℃灭菌5 min)、酵母膏、琼脂粉 广东环凯微生物科技有限公司;胰蛋白胨、乳糖、半乳糖、蛋白胨、果糖、蔗糖、大豆蛋白胨、浓硫酸、无水乙醇 国药集团化学试剂有限公司;1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH) 北京陆桥技术有限责任公司;酚酞 天津市化学试剂研究所;三氯乙酸 上海真能实业有限公司;试剂均为分析纯。
1.2 仪器与设备
GZ-400-S生化培养箱 韶关市广智科技设备有限公司;LDZX-50KBS立式压力蒸汽灭菌器 上海申安医疗器械厂;JA2003电子天平 上海浦春计量仪器有限公司;SW-CJ-2D净化工作台 苏州博莱尔净化设备有限公司;101-AB pH计 天津市泰斯特仪器有限公司;RW-20D-S25超级恒温水浴锅 德国IKA公司;BCD-328WDPT紫外分光光度计 青岛海尔股份有限公司;QP2010台式高速离心机 岛津企业管理(中国)有限公司。
1.3 方法
1.3.1 菌种活化
取―80 ℃保藏的菌种按体积分数5%(下同)接种量接种于MRS液体培养基,37 ℃培养24 h,活化2 代后备用。
1.3.2 单因素试验设计
1.3.2.1 最适培养基初始pH值确定
参照萨如拉[10]的方法,将植物乳杆菌XN1904E种子液按5%接种量接种于MRS液体培养基,用0.1 mol/L NaOH和0.1 mol/L HCl调节培养基起始pH值,分别调节至pH 6.0、6.5、7.0、7.5、8.0,在37 ℃条件下发酵培养24 h,测定发酵液中胞外多糖含量,确定最适培养基初始pH值。
1.3.2.2 最适发酵温度确定
在最适pH值条件下,将植物乳杆菌XN1904E种子液按5%接种量接种于MRS液体培养基,设置培养温度分别为35、37、39、41、43 ℃,培养24 h,测定发酵液中胞外多糖含量,确定最适发酵温度[11]。
1.3.2.3 最适碳源确定
以MRS培养基为基础培养基,培养基中的葡萄糖分别用等质量蔗糖、乳糖、半乳糖和果糖替换,按5%接种量接种活化后植物乳杆菌XN1904E到培养基中,在最适pH值和最适发酵温度下培养24 h,测定发酵液中胞外多糖含量,确定最适碳源。
1.3.2.4 最适氮源确定
以MRS培养基为基础培养基,培养基中的蛋白胨分别用等质量鱼蛋白胨、胰蛋白胨、酵母膏替换,按5%接种量接种活化后植物乳杆菌XN1904E到添加最适碳源的培养基中,在最适pH值和最适发酵温度下培养24 h,测定发酵液中胞外多糖含量,确定最适氮源。
1.3.3 正交试验设计
根据上述单因素试验结果,选择影响较为显著的因素及水平,进行4因素3水平正交试验优化[12]。各因素及水平如表1所示。
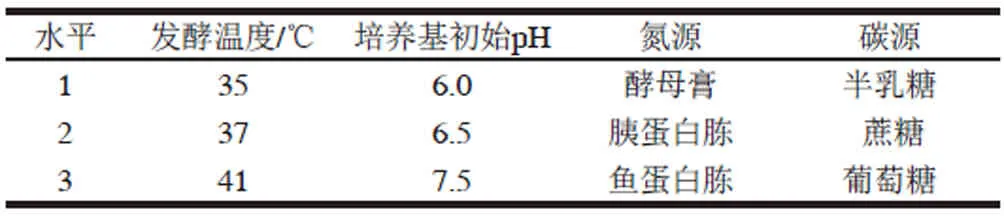
表 1 正交试验因素水平表Table 1 Level for independent variables used in orthogonal array design
1.3.4 植物乳杆菌XN1904E胞外多糖含量测定
粗胞外多糖的提取:参照黄承敏等[1]方法,将纯化得到的植物乳杆菌XN1904E接种到MRS液体培养基培养18 h,再按5%接种量接种到MRS液体培养基,37 ℃培养24 h。取10 mL发酵液10 000×g离心10 min,去菌体,上清液于沸水浴中加热10 min,冷却至室温;加入2 倍体积10 g/100 mL三氯乙酸溶液,室温下充分搅拌,静置30 min,10 000×g、4 ℃离心15 min,上清液装入透析袋(截留分子质量14 000 Da),4 ℃冰箱中超纯水透析24 h;加入3 倍体积体积分数95%乙醇,4 ℃过夜,10 000×g、4 ℃离心15 min,去上清液,沉淀溶于10 mL蒸馏水,装入透析袋,透析2 d,每8 h换1 次水,得胞外多糖水溶液。
胞外多糖含量测定:准确称取10 mg烘干至恒质量的葡萄糖标准品,置于100 mL容量瓶中,蒸馏水定容;分别移取0、3、6、9、12、15 mL上述标准溶液稀释,制得质量浓度分别为0、30、60、90、120、150 μg/mL的葡萄糖标准溶液,分别移取不同质量浓度葡萄糖标准溶液1 mL于比色管中,加入1 mL质量分数6%苯酚溶液,再加入4 mL浓硫酸摇匀,室温下静置30 min,于490 nm波长处测定吸光度。以吸光度为横坐标,葡萄糖标准溶液质量浓度为纵坐标,绘制标准曲线,所得标准曲线方程为y=0.006 4x+0.011 6(R2=0.999 4)。取1 mL胞外多糖水溶液,加入1 mL质量分数6%苯酚溶液,再加入4 mL浓硫酸,摇匀,室温下静置30 min,于490 nm波长处测定吸光度,以MRS液体培养基为对照,最后按葡萄糖标准曲线方程计算胞外多糖含量[13]。
1.3.5 植物乳杆菌XN1904E胞外多糖水溶液DPPH自由基清除率测定
0.2 mmol/L DPPH乙醇溶液的配制:准确称取19.7 mg DPPH,溶于无水乙醇,用250 mL容量瓶定容,避光保存于4 ℃冰箱备用[14]。
取1 mL胞外多糖水溶液,注入洁净试管中,以1 mg/mL VC作为阳性对照,分别加入1 mL 0.2 mmol/L DPPH乙醇溶液混匀,室温下避光反应30 min,用分光光度计测定517 nm波长处吸光度(Ai);空白组以等体积乙醇溶液代替DPPH乙醇溶液,测定517 nm波长处吸光度(Aj);对照组以等体积蒸馏水代替样品溶液,测定517 nm波长处吸光度(A0)。DPPH自由基清除率按下式计算。每组做3 个平行。
1.4 数据处理
结果用平均值±标准差表示,实验数据采用Excel 2010、Origin 2018和DPS数据统计软件进行处理,用t-检验进行分析,P<0.05为差异显著。
2 结果与分析
2.1 单因素试验结果
2.1.1 培养基初始pH值对植物乳杆菌XN1904E产胞外多糖的影响培养基初始pH值是微生物生长环境的重要指标,通过改变酶活性影响微生物新陈代谢。控制培养基初始pH值是提高乳酸菌胞外多糖产量的重要条件[15]。由图1可知,在不同培养基初始pH值条件下,植物乳杆菌XN1904E的胞外多糖产量也不同。当培养基初始pH值为6.5时,胞外多糖产量达到最大,为(163.16±8.82) mg/L。在偏弱酸性条件下,胞外多糖产量高于碱性条件,植物乳杆菌XN1904E产胞外多糖的最适初始pH值为6.5。此结果说明乳酸菌胞外多糖产量与培养基初始pH值有依赖关系,过高或过低的pH值都会影响菌种生长,从而影响胞外多糖产量。
2.1.2 发酵温度对植物乳杆菌XN1904E产胞外多糖的影响温度通过影响蛋白质、核酸等生物大分子结构及胞内酶活性等,影响微生物生长[16]。由于微生物多糖属于代谢产物,发酵温度过低导致乳酸菌菌体生长缓慢,影响代谢产物胞外多糖的含量[17]。由图2可知,在37 ℃条件下,胞外多糖产量最高,达(196.46±10.92) mg/L,随着发酵温度升高,植物乳杆菌XN1904E的胞外多糖产量呈现先升高后降低的趋势(P<0.05)。这可能是由于发酵温度过高,稳定期提前到来,使得生物酶失去活性,导致多糖产量降低,植物乳杆菌XN1904E产胞外多糖的最适发酵温度为37 ℃。
2.1.3 碳源对植物乳杆菌XN1904E产胞外多糖的影响
碳源是微生物生长代谢重要的营养物之一,主要提供细胞合成产物的碳架及代谢所需能量,由于不同微生物的碳水化合物代谢酶系不同,因此其可利用的碳源也不同[18]。由图3可知,半乳糖作为碳源时,植物乳杆菌XN1904E胞外多糖产量最高,达到(190.89±6.06) mg/L,其次果糖作为碳源时, 胞外多糖产量为(189.29±8.32) mg/L,蔗糖和葡萄糖作为碳源时胞外多糖产量分别为(165.56±15.21)、(150.22±2.84) mg/L,乳糖作为碳源时胞外多糖产量最低。半乳糖最适合作为植物乳杆菌XN1904E产胞外多糖的碳源。
2.1.4 氮源对植物乳杆菌XN1904E产胞外多糖的影响氮源的主要作用是为微生物细胞生长和代谢提供氮素及能量[19]。由图4可知,鱼蛋白胨作为氮源时,植物乳杆菌X N 1 9 0 4 E 的胞外多糖产量最高,达(196.36±6.25) mg/L,胰蛋白胨和酵母膏作为氮源时胞外多糖产量分别为(152.22±4.37)、(126.09±7.46) mg/L,蛋白胨作为氮源时植物乳杆菌XN1904E胞外多糖产量最低,为(106.49±5.26) mg/L,所以植物乳杆菌XN1904E的最适氮源为鱼蛋白胨。鱼蛋白胨比胰蛋白胨和普通蛋白胨更有利于植物乳杆菌XN1904E产生胞外多糖,这可能与各蛋白胨的组成差异有关,说明优质的蛋白源更有利于产生胞外多糖。
2.2 植物乳杆菌XN1904E培养条件优化结果
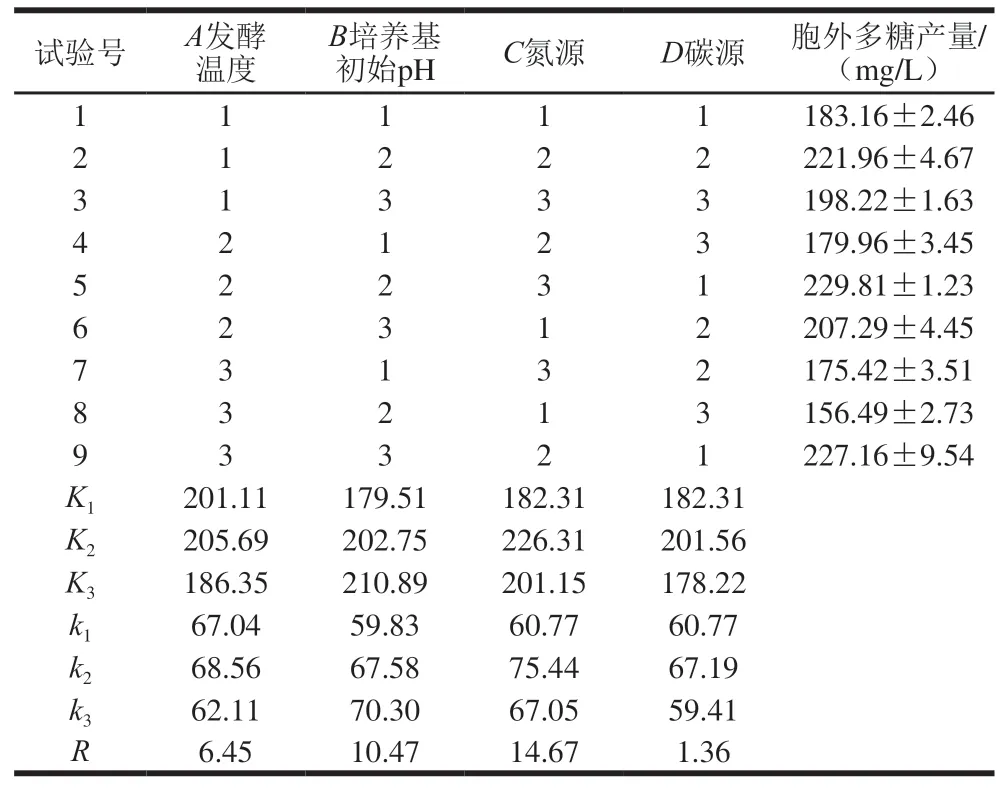
表 2 正交试验结果Table 2 Orthogonal array design with experimental results
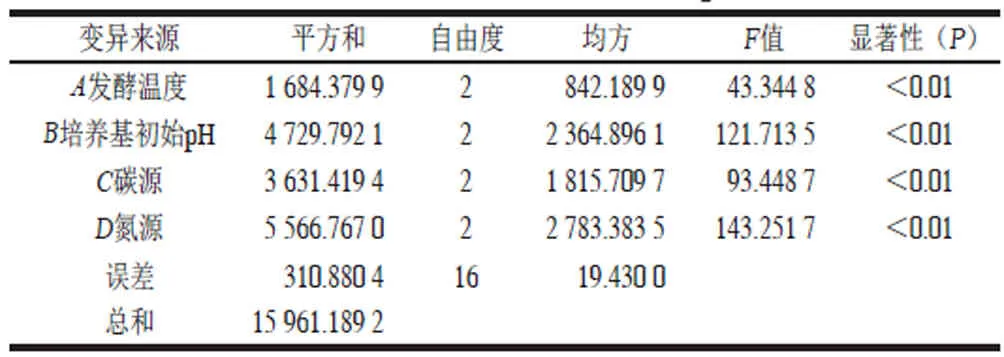
表 3 正交试验结果方差分析Table 3 Analysis of variance for the effect of fermentation conditions on exopolysaccharide production by Lactobacillus plantarum XN1904E
由表2可知,试验影响因素的主次顺序为C>B>A>D,即氮源>培养基初始pH值>发酵温度>碳源。由表3可知,方差结果显示,发酵温度、培养基初始pH值、碳源和氮源对胞外多糖产量均影响极显著(P<0.01)。试验最优方案为A2B2C3D1,即最佳培养条件为发酵温度37 ℃、培养基初始pH 6.5、氮源选择鱼蛋白胨、碳源选择半乳糖,此时胞外多糖产量为(229.81±1.23) mg/L。
2.3 植物乳杆菌XN1904E胞外多糖水溶液DPPH自由基清除率
DPPH自由基清除能力可作为评判抗氧化能力强弱的指标,是维持人体内自由基平衡的重要因素之一[20]。为初步探索植物乳杆菌XN1904E所产胞外多糖是否具有抗氧化能力,对该胞外多糖水溶液进行DPPH自由基清除率测定。结果表明,植物乳杆菌XN1904E胞外多糖水溶液(0.2 mg/mL)的DPPH自由基清除率为27.54%。Rajoka等[21]从母乳中分离得到的乳酸菌胞外多糖质量浓度为0.4 mg/mL时,自由基清除率最高,为30%左右。本研究与此结果略有偏差,可能是因为菌株的不同,但能初步判定植物乳杆菌XN1904E所产胞外多糖具有抗氧化能力,其他抗氧化能力还需进一步研究。
3 结 论
通过对植物乳杆菌XN1904E培养条件进行优化发现,产胞外多糖最适初始pH值为6.5,最适发酵温度为37 ℃,最适碳源为半乳糖,最适氮源为鱼蛋白胨;在单因素试验基础上通过正交试验得出植物乳杆菌XN1904E的最适培养条件为初始pH 6.5、发酵温度37 ℃、半乳糖最适碳源,鱼蛋白胨为最适氮源。通过验证实验,测得最适培养条件下植物乳杆菌XN1904E胞外多糖产量为235.41 mg/L。植物乳杆菌XN1904E胞外多糖水溶液(0.2 mg/mL)对DPPH自由基的清除率为27.54%,表明其具有一定的抗氧化能力。
猜你喜欢
油气田地面工程(2022年9期)2022-10-26
江苏农业科学(2022年6期)2022-04-15
铀矿冶(2022年1期)2022-02-17
中老年保健(2021年9期)2021-08-24
昆钢科技(2021年6期)2021-03-09
农家顾问(2016年12期)2017-01-06
药学研究(2015年11期)2015-12-19
分析化学(2014年10期)2014-10-24
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年13期)2014-03-11
