
菊芋全粉对高温胁迫下仿刺参免疫及消化功能的影响
2020-06-29 03:38朱晓振孟现尧贾晨晨于殿江张美超衣悦涛
中国饲料 2020年11期
朱晓振,孟现尧,贾晨晨,于殿江,张美超,衣悦涛
(1.中国科学院烟台海岸带研究所,山东烟台 264003;2.中国科学院大学,北京 100049;3.烟台大学海洋学院,山东烟台 264005)
仿刺参(Apostichopus japonicas)属于棘皮动物门(Echinodermata),海参纲(Holothurooidea),是我国北方水产养殖中的重要经济物种之一 (李成林等,2010)。仿刺参具有丰富的营养组成,同时也具有很高的药用价值和经济价值,自古以来就享有“海八珍”的美称。但近年来受气候变化影响,夏季持续高温和降雨,对刺参池塘养殖造成严重影响。高温导致水体溶解氧降低,影响仿刺参生理行为及代谢,同时会导致厌氧细菌大量繁殖并对有机物进行厌氧分解,产生大量氨氮、硫化氢、亚硝酸盐等有害物质,损伤仿刺参的非特异性免疫系统,增加对病原菌的易感性(霍达等,2017;徐松涛等,2017)。另外,持续性的高温会导致刺参出现消化道萎缩,消化功能减弱,体重减轻的现象。当环境温度发生变化时,仿刺参肠道中的消化酶活性也会发生相应的改变,在适宜的温度范围内,适当的温度升高会提高仿刺参肠道内消化酶的活力,对消化能力起到促进的作用,但一旦突破了临界温度,消化酶活性就会受到抑制甚至失活,进而对仿刺参的消化能力与生长产生影响,严重时会出现大面积死亡(杨秀生等,2009)。
益生元是一种不可被消化的食物组成部分,一些功能性低聚糖类、多糖类、天然植物提取物、蛋白质水解物、多元醇等均属于益生元。益生元的结构特点为: 能选择性地刺激肠道中有益菌群的生长繁殖和激活其代谢功能;使肠道菌群向有利于寄主健康的方向转化;刺激免疫,提高机体免疫力;改善肠道内环境,还富有多种营养物质,促进生长。益生元作为一种新型绿色功能性饲料添加剂逐渐成为了动物营养与研究的新方向,已有各种各样的益生元作为饲料添加剂应用于畜禽、水产的养殖生产中(邓紫艳等,2015)。菊芋全粉作为一种富含益生元的植物多糖,其主要成分菊粉是一种水溶性果聚糖益生元,可调节动物机体肠道微生物组成、改善肠胃功能、增强免疫功能,被广泛应用于食品和动物饲料中(陈佳亿等,2018)。本试验旨在探究菊芋全粉作为仿刺参饲料添加剂对高温胁迫下仿刺参的消化能力及免疫力的影响,为生产实践提供理论依据。
1 材料与方法
1.1 试验动物 试验用仿刺参选用山东东方海洋科技股份有限公司育苗车间1 龄以上的刺参。挑选质量与活力较好的仿刺参,分别暂养于8 个容量为40 L 的养殖水箱中,每个水箱中随机投放50 头仿刺参。每日下午15 点定时投喂饲料,饱食投喂,投喂量一般为仿刺参体重的3%,投喂前及时清理残饵及粪便,换水量为养殖水箱总水体的1/3。暂养1 周后用于试验。
1.2 试验分组 暂养结束后,选择体格健壮、质量较好的仿刺参320 头,随机分为8 组,饲喂基础饲料一个月;一个月后设置4 个温度梯度,分别为18、23、28、32 ℃,每个温度下设置 2 组,其中基础组投喂基础饲料,试验组投喂添加15‰菊芋全粉的试验饲料(分组情况见表1),暂养7 d,能正常摄食后进行分组试验,试验开始时称重一次并记录。

表1 仿刺参分组情况
1.3 高温胁迫试验 基础温度为18 ℃,升温幅度为1 ℃(24 h),达到试验温度后进行恒温饲养,当所有试验组都到达预定温度后,再按照其当前温度恒温饲养两周,在高温胁迫下将基础饲料和添加菊芋全粉的饲料喂养的仿刺参进行对比研究,观察仿刺参生长状态,记录成活率,高温胁迫结束后,取仿刺参体腔液和肠道样品,测定仿刺参体腔液中的各种免疫酶活性和消化酶活性。
1.4 测定指标与方法
1.4.1 成活率测定 分别对不同试验组仿刺参每日的死亡情况进行记录,饲养阶段结束后,计算不同试验组仿刺参的存活率(SR)。
SR/%=Ni/No×100%。
式中:Ni 是饲养结束后仿刺参数量,No 是饲养开始时仿刺参数量。
1.4.2 免疫因子活力测定 在两周高温胁迫试验结束后,分别从每组取出9 只健康仿刺参在冰盘上取体腔液样品,于 4 ℃、3000×g 离心 10 min,取上清液保存在-80 ℃冰箱中备用,用作体腔液免疫指标的测定。体腔液样品中过氧化氢酶(CAT)、酸性磷酸酶(ACP)、碱性磷酸酶(AKP)、溶菌酶(LSZ)、一氧化氮合酶(NOS)活性均采用试剂盒(南京建成生物科技有限公司)测定。
1.4.3 消化酶活力的测定 在两周高温胁迫试验结束后,分别从每组取出9 只健康仿刺参在冰盘上取肠道样品,并且除去仿刺参肠道上黏附的呼吸树及其他不相干成分,用无菌滤纸去除多余水分,置于冻存管内,-80 ℃下保存,用于消化酶的测定。测定前,将冷冻的肠道样品取出解冻,准确称取待测肠道重量,加入9 倍体积生理盐水,在4 ℃的冰水浴条件下进行匀浆。匀浆结束后将样品2500×g 离心10 min,然后取上清液放入冻存管待用。蛋白酶测定采用福林-酚法,淀粉酶测定采用碘-淀粉比色法,脂肪酶测定采用滴定法(樊英等,2017)。仿刺参肠道样品蛋白酶、脂肪酶、淀粉酶活性采用试剂盒(南京建成生物科技有限公司)测定。
1.5 数据处理与分析 数据用SPSS 16.0 软件进行单素方差分析 (One-way AN0VA),结果以“平均值±标准差”表示,当P <0.05 时为差异显著。
2 结果与分析
2.1 菊芋全粉对高温胁迫仿刺参成活率的影响由图1 可知,在18 ℃及23 ℃的生长环境下,菊芋全粉组相较于普通饲料组,在存活率方面表现的更优。当温度达到28 ℃,菊芋全粉组仿刺参成活率 (88.75±1.25)%明显高于普通饲料组 (85.25±2.75%,P < 0.05)。这说明通过添加 15 g/kg 菊芋全粉能在一定程度上改善高温胁迫下仿刺参的成活率。32 ℃时,菊芋全粉组成活率显著下降,机理需要进一步研究。
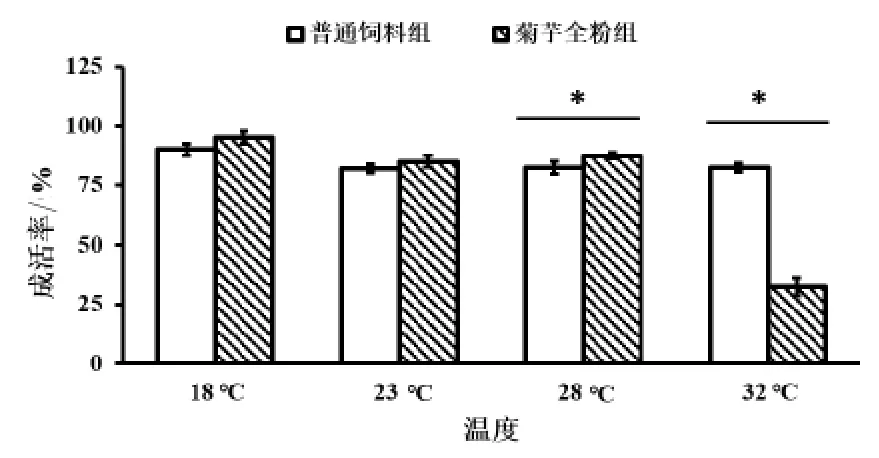
图1 菊芋全粉对高温胁迫下仿刺参成活率的影响
2.2 菊芋全粉对高温胁迫下仿刺参免疫因子变化的影响
2.2.1 菊芋全粉对仿刺参体液中过氧化氢酶(CAT)活力的影响 由图2 可知,在同一温度下,普通饲料组和菊芋全粉组CAT 活力差异不显著。随着温度升高,与18 ℃和23 ℃相比,在28 ℃和32 ℃刺激下,仿刺参 CAT 活力显著升高(P < 0.05),其中普通饲料组和菊芋全粉组没有显著性差异。

图2 菊芋全粉对仿刺参体液中过氧化氢酶(CAT)活力的影响
2.2.2 菊芋全粉对仿刺参体液中酸性磷酸酶(ACP)活力的影响 由图3 可知,在 18 ℃和 28 ℃温度下,相较于普通饲料组 (2.74±0.26、0.18±0.19 U/100 mL),菊芋全粉组 (3.11±0.53、2.59±0.29 U/100 mL)的仿刺参ACP 活力明显提高 (P <0.05),23 ℃和32 ℃干预下,普通饲料组和菊芋全粉组仿刺参的ACP 活力没有显著性差异。
2.2.3 菊芋全粉对仿刺参体液中碱性磷酸酶(AKP)活力的影响 由图4 可知,在同一温度下,菊芋全粉组和普通饲料组无显著差异;28 ℃菊芋全粉组(8.86±0.20 U/100 mL)与 18 ℃和 23 ℃菊芋全粉组(7.82±0.24、8.39±0.47 U/100 mL)相比具有显著性差异(P <0.05)。普通饲料组和菊芋全粉组的AKP 活力均呈现先增长后下降的趋势,但是各个试验组的差异并不显著,其中28 ℃菊芋全粉组AKP 活力最高。

图3 菊芋全粉对仿刺参体液中酸性磷酸酶(ACP)活力的影响
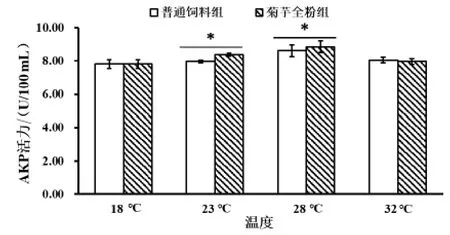
图4 菊芋全粉对仿刺参体液中碱性磷酸酶(AKP)活力的影响
2.2.4 菊芋全粉对仿刺参体液中溶菌酶(LSZ)含量的影响 由图5 可知,随着胁迫温度的升高,普通饲料组与菊芋全粉组的仿刺参体液中LSZ 含量都处于上升趋势。而且28 ℃普通饲料组 (1.72±0.53 μg/mL)与菊芋全粉组(2.82±0.50 μg/mL) LSZ活力具有显著性差异(P < 0.05)。32 ℃干预下,与各组相比,菊芋全粉组LSZ 活力呈现最大值。

图5 菊芋全粉对仿刺参体腔液中溶菌酶(LSZ)含量的影响
2.2.5 菊芋全粉对仿刺参体液中一氧化氮合酶(NOS) 活力的影响 由图6 可知,在 18 ℃条件下,与普通饲料组相比,菊芋全粉组NOS 活力升高,说明在正常仿刺参养殖环境下,通过在饲料中添加菊芋全粉,可以改善NOS 活力。23 ℃普通饲料组NOS 活力显著高于32 ℃普通饲料组;其次,在 18 ℃和 32 ℃条件下,菊芋全粉组(5.41±0.21、4.71±0.80 U/mL)NOS 活力明显高于普通饲料组(4.90±0.79、3.79±0.67 U/mL);而在 23 ℃条件下普通饲料组与菊芋全粉组差别很大。随着温度升高,普通饲料组NOS 活力呈现先增高后降低的趋势,添加菊芋全粉组呈现先降低后升高趋势。
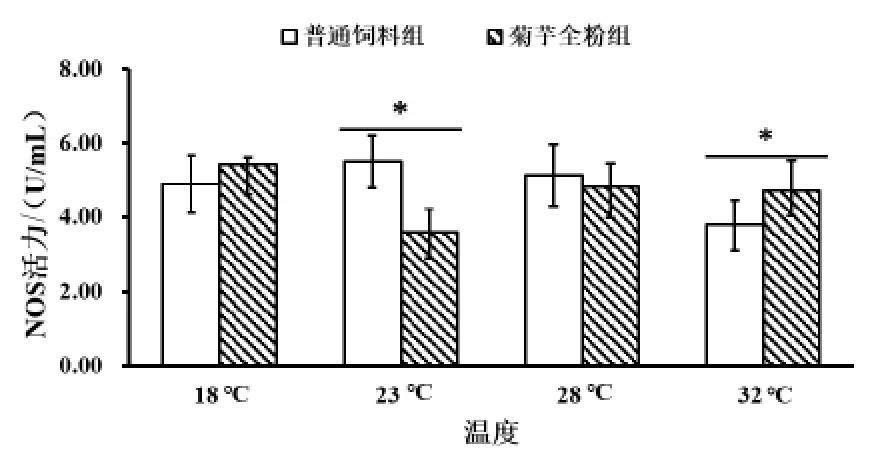
图6 菊芋全粉对仿刺参体腔液中一氧化氮合酶(NOS)活力的影响
2.3 菊芋全粉对高温胁迫下仿刺参消化酶活力的影响
2.3.1 菊芋全粉对对高温胁迫下仿刺参肠道蛋白酶活力的影响 由图7 可知,随着温度的升高,普通饲料组和菊芋全粉组蛋白酶活力均升高,普通饲料组 (72.22±12.64 U/mL) 与菊 芋全 粉组(58.47±13.54 U/mL) 在 32 ℃时有显著差异,并且蛋白酶活力明显高于其他温度组,其他温度下添加菊芋全粉的饲料对蛋白酶活力没有显著影响。
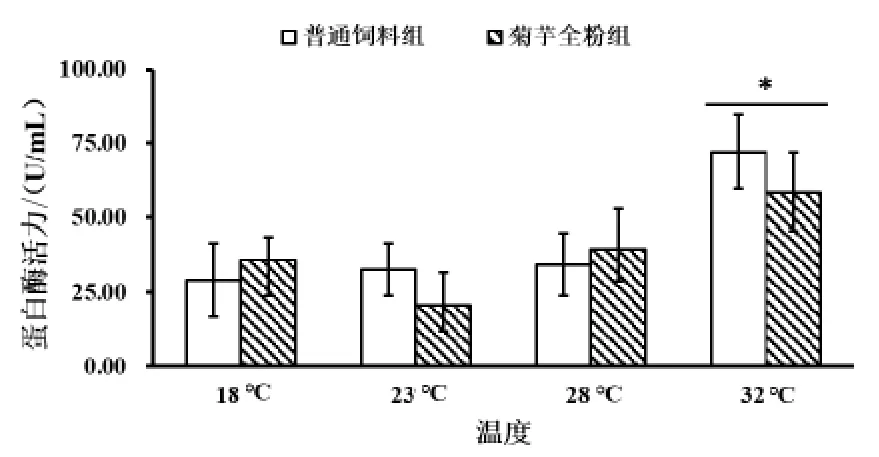
图7 菊芋全粉对高温胁迫下仿刺参肠道蛋白酶活力的影响
2.3.2 菊芋全粉对高温胁迫下仿刺参肠道脂肪酶活力的影响 由图8 可知,随着温度升高,仿刺参肠道脂肪酶活力下降,在28 ℃时,菊芋全粉组(2.82±0.50 U/mL)脂肪酶活力显著高于普通饲料组(1772±0.53 U/mL;P < 0.05)。
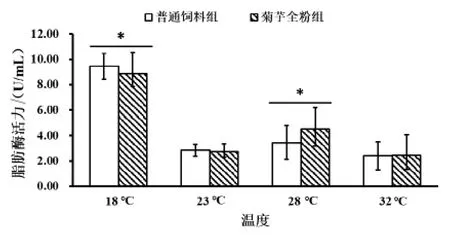
图8 菊芋全粉对高温胁迫下仿刺参肠道脂肪酶活力的影响
2.3.3 菊芋全粉对高温胁迫下仿刺参肠道淀粉酶活力的影响 由图9 可知,在普通饲料组和菊芋全粉组中淀粉酶活力随着温度升高而降低,相比18 ℃,当温度升高至32 ℃时,与菊芋全粉组(0.20±0.08 U/mL) 相比,普通饲料组 (0.10±0.02 U/mL) 淀粉酶活力下降更显著 (P < 0.05)。在 18 ℃与 32 ℃时,普通饲料组 (0.37±0.05、0.10±0.02 U/mL) 与菊芋全粉组 (0.31±0.05、0.20±0.08 U/mL) 的淀粉酶活力在组内存在显著差异(P < 0.05),其他试验组变化不显著。在高温胁迫下,通过在饲料中添加菊芋全粉,有减缓高温胁迫下淀粉酶活力降低的作用。
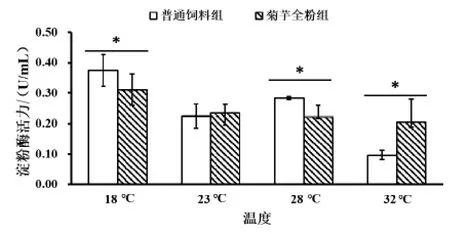
图9 菊芋全粉对高温胁迫下仿刺参肠道淀粉酶活力的影响
3 讨论
3.1 菊芋全粉对仿刺参成活率的影响 益生元在水产养殖业中的作用越来越受到重视,研究表明益生元不仅可改善水产动物的生长性能(Guerreiro 等,2015),还可有效降低患病率和死亡率。本研究表明,在18、23 ℃和28 ℃温度下,菊芋全粉组成活率明显高于普通饲料组,说明在高温胁迫下通过添加菊芋全粉可以降低仿刺参死亡率,添加菊芋全粉在一定高温范围内可以增强仿刺参在高温胁迫下抵抗疾病和应激的能力,促使其成活率提高。有研究发现,在杂交罗非鱼养殖过程中,在饲料中添加不同比例的甘露寡糖和低聚果糖可以提高动物成活率,增强非特异性免疫(何四旺等,2003)。温度超过 32 ℃时,菊粉全粉组与普通饲料组成活率没有差异,这时温度已经超过刺参的耐受温度值,导致仿刺参死亡。仿刺参随体重的增加对高温的耐受逐渐下降,小于25 g/头的稚参在28 ℃仍可正常活动,而体重在160 g/头左右的仿刺参在 22 ℃左右会进入夏眠状态。此时仿刺参停止摄食,代谢水平较低,当水温超过夏眠承受温度时,仿刺参会出现吐肠、化皮等应激反应;当水温升至 31 ℃以上时,会化皮死亡 (贺加贝等,2019)。
3.2 菊芋全粉对仿刺参免疫因子变化的影响温度是影响仿刺参生长和发育的重要因素,夏季高温,养殖水体中硫化物及氨氮含量超标,影响仿刺参存活,破坏仿刺参机体非特异性免疫防御系统,降低免疫力,导致对病原菌的易感性增加(徐松涛等,2017;臧元奇等,2012)。益生元作为功能性成分,目前已有较多研究表明其可改善水产动物肠道健康,增强机体免疫力,高温胁迫环境中体内产生的大量活性氧会造成机体氧化损伤和免疫机能破坏,此时机体抗氧化防御酶系统(如 CAT等)启动免疫应答。研究发现,在温度由16 ℃上升至 26 ℃时,仿刺参机体的CAT 活力上升,持续26 ℃胁迫下又回落至较低水平(田相利等,2014;Ji 等,2008)。本研究中普通饲料组与菊芋全粉组CAT 活力都随着温度的升高而升高,在28 ℃高温胁迫下,菊芋全粉组 (16.15±0.96 U/mL) CAT 活力高于普通饲料组 (15.81±0.53 U/mL),但没有显著性差异。ACP 和AKP 是两种重要水解酶,可催化各种含磷化合物的水解过程。ACP 是巨噬细胞溶酶体的标志酶,其活性反映吞噬细胞清除异物的能力(刘云等,2008)。在 18 ℃和 28 ℃温度下,菊芋全粉组 (3.11±0.53、2.59±0.29 U/100 mL) 的ACP 活力明显提高。说明菊芋全粉在一定程度上可以缓解高温胁迫下对仿刺参造成的机体免疫力下降。随着胁迫温度的升高和时间的延长,普通饲料组和菊芋全粉组的 ACP 活力呈现降低趋势,研究结果显示菊芋全粉可抑制ACP 活力降低的程度,这与刺参在干露和夏眠等胁迫条件下酶活力变化基本一致 (田相利等,2014;Du 等,2013)。AKP 是一种对底物专一性要求较低的磷酸单脂水解酶,可以起到解毒的功能,有改善机体免疫力的作用(Zhang 等,2000)。本研究中,AKP 活力在普通饲料组和菊芋全粉组均呈现先升后降低的趋势,在胁迫温度达到28 ℃时,菊芋全粉组 (8.86±0.20 U/100 mL) AKP 活力显著高于普通饲料组(8.62±0.37 U/100 mL),同时当温度高于 28 ℃时,菊芋全粉组可以减缓AKP 活力降低程度。说明在高温胁迫下,通过在饲料中添加菊芋全粉可以使仿刺参保持较强的免疫防御能力。LSZ 是吞噬细胞杀菌的物质基础,是非特异性免疫的重要组成部分,担负着机体防御的功能。研究发现随着温度升高,LSZ 活力会升高,温度超过 32 ℃,动物机体免疫系统受损,LSZ 活力也逐渐降低;同样有研究发现在 32 ℃下处理 6~12 h 的仿刺参体腔液LSZ 活力显著降低(Wang 等,2008)。在本研究中,随着温度升高,普通饲料组和菊芋全粉组LSZ 活力均升高,且在28℃高温胁迫下,菊芋全粉组(2.82±0.50 μg/mL) LSZ 活力具有显著性差异,说明在高温刺激下,菊芋全粉可以增强刺参机体LSZ 活力,使得刺参在高温胁迫能够更有效地做出免疫调节反应,保护机体免受损伤。NOS 可以催化底物中L -精氨酸,在动物机体内产生NO,NO 可杀伤病毒、细菌,抑制感染,在免疫中起着重要的作用,NOS 活性影响NO 含量,从而体现免疫应激反应的情况(王广军等,2005)。随着温度升高,普通饲料组NOS 活力明显下降,32 ℃时下降最明显(3.79±0.67 U/mL);相反,菊芋全粉的添加,降低了NOS 活力下降程度,在32 ℃高温胁迫下,菊芋全粉组 (4.71±0.80 U/mL) NOS 活力显著高于普通饲料组 (3.79±0.67 U/mL)。这说明,通过在饲料中添加菊芋全粉,使得仿刺参表现出对高温胁迫的较强适应性。通过在饲料中添加菊芋全粉,在高温胁迫下,能一定程度上增强刺参免疫因子活力,使得仿刺参在高温胁迫下具有更好的耐受能力。
3.3 菊芋全粉对仿刺参消化酶活性的影响 消化酶是机体多种酶中的一部分,仿刺参肠道消化酶种类很多,主要是蛋白酶、淀粉酶、脂肪酶等。仿刺参为低温性变温物种,环境温度是影响酶活性的重要因素,在夏眠过程中,仿刺参体内与代谢和免疫相关的酶活性会发生变化。消化酶(淀粉酶、脂肪酶)活性会下降,只有蛋白酶在此期间显著升高(任庆印等,2013)。本研究中,随着温度升高,普通饲料组和菊芋全粉组蛋白酶活性均升高,淀粉酶活力和脂肪酶活力均下降。研究发现,仿刺参蛋白质含量丰富,需要依赖高活性蛋白酶摄取食物中的蛋白质,以合成自身蛋白,因此在刺参中蛋白酶活力最高(姜令绪等,2007)。在高温胁迫下,32 ℃时普通饲料组 (72.22±12.64 U/mL) 蛋白酶活性显著升高,通过在饲料中添加菊芋全粉(58.47±13.54 U/mL),为维持机体稳定,高温胁迫下降低了蛋白酶活性的升高趋势。脂肪酶与淀粉酶同属内源性消化酶,在刺参消化酶中活力最低(姜令绪等,2007)。随着温度升高,普通饲料组和菊芋全粉组脂肪酶活性都降低,菊芋全粉对脂肪酶活力没有显著影响。消化酶活性与仿刺参的营养吸收和生长关联密切,消化酶活性的降低,影响着仿刺参的生长状态与成活率。仿刺参肠道中淀粉酶也是主要的消化酶,在消化道不同部位的活性不同(袁成玉等,2007)。随着胁迫温度的升高,普通饲料组淀粉酶活性显著降低,相反,菊芋全粉明显降低了淀粉酶活性降低趋势,说明在高温胁迫下,添加菊芋全粉可维持仿刺参淀粉酶活性相对稳定。研究表明,在饲料中添加益生元能够增加肠道褶皱、肠上皮细胞以及微绒毛的高度,进而改变胃肠道的吸收区(Dimitroglou 等,2010)。在高温胁迫下,通过补充菊芋全粉,淀粉酶活力降低趋势得到改善,使得动物处于应激状态时表现得更为突出。
4 结论
与普通饲料相比,高温胁迫下,在饲料中添加菊芋全粉,在28 ℃以下,可以显著提高仿刺参成活率;高温胁迫下仿刺参免疫因子ACP 和AKP在菊芋全粉的影响下,可以维持相对稳定,随着胁迫温度的升高,菊芋全粉组LSZ 活力显著升高;高温胁迫下,菊芋全粉对蛋白酶和脂肪酶的影响不大,但是可以显著降低高温刺激下淀粉酶活力降低趋势。本试验条件下,在饲料中添加15 g/kg菊芋全粉,在高温胁迫下对于仿刺参成活率、免疫因子以及消化酶活性有一定改善作用。
猜你喜欢
现代面粉工业(2022年2期)2022-12-08
中国马铃薯(2022年4期)2022-10-18
中国马铃薯(2022年3期)2022-09-13
红蜻蜓·低年级(2021年12期)2022-01-19
红蜻蜓·低年级(2021年12期)2021-12-19
保鲜与加工(2021年11期)2021-11-26
活力(2021年6期)2021-08-05
渔业科学进展(2021年3期)2021-05-12
人大建设(2020年1期)2020-11-17
大连海洋大学学报(2020年2期)2020-05-06
