
超极化激活环核苷酸门控阳离子通道亚型在大鼠脊髓背角浅层的特异性表达*
2020-06-09 02:41黄姣艳程小娥马龙先张达颖蒋昌宇涛3
中国病理生理杂志 2020年5期
黄姣艳, 程小娥, 马龙先, 张达颖, 蒋昌宇, 柳 涛3,△
(南昌大学第一附属医院 1麻醉科,2疼痛科,3医学科研中心,江西南昌330006;4深圳市南山医院韩济生院士疼痛医学工作站,广东深圳518052)
超级化激活环核苷酸门控阳离子通道(hyperpolarization-activated cyclic nucleotide-gated cation channels,HCN channels)有HCN1~4共4种亚型,主要表达在神经细胞和心肌细胞的胞膜上,与癫痫、抑郁、疼痛以及心脏疾病的发生发展有着密切的关联[1-4]。脊髓背角(spinal dorsal horn,SDH)是外周伤害性信息向中枢传递的初级门户,该区域的HCN通道已被证实在慢性疼痛的发生发展中具有重要的作用。例如大鼠脊髓背角HCN2的表达在慢性炎性痛后显著增加,而阻断或抑制HCN通道可大幅减轻神经病理性疼痛和炎性痛模型动物的痛行为[5-7]。SDH由浅至深分为Ⅰ~Ⅵ层,其中Ⅰ~Ⅱ层构成脊髓背角浅层,对疼痛的传递与整合起着重要的调控作用。我们近期研究初步表明,HCN通道在脊髓背角浅层广泛表达,且HCN4与兴奋性轴突末梢标志物囊泡谷氨酸转运体2(vesicular glutamate transporter 2,VGLUT2)显著共染[8-9],但HCN通道在神经元和胶质细胞中的具体表达情况及在神经元亚细胞结构如树突、轴突等部位的分布尚不清楚。因此,本研究应用免疫组织化学和激光共聚焦成像技术,观察SDH中HCN通道的4种亚型在正常大鼠脊髓背角浅层的特异性分布情况,以期为进一步揭示SDH的HCN通道在痛信号通路中的作用提供参考资料。
材料和方法
1 动物
3~5周龄清洁级 Sprague-Dawley(SD)大鼠 27只,体重80~100 g,雌雄不拘,来源于南昌大学动物中心[许可证号为SYXK(赣)2005-0001]。动物实验严格遵循《南昌大学实验动物管理办法》和《南昌大学动物实验伦理审查》的原则进行。实验大鼠饲养于清洁笼具中,室内光照保持12 h/12 h昼夜循环,室温控制在20~25℃,相对湿度在65%~70%之间,自由饮食。
2 主要试剂及仪器
抗HCN1~4抗体购自Alomone Labs;抗胶质细胞原纤维酸性蛋白(glial fibrillary acidic protein,GFAP)抗体、抗降钙素基因相关肽(calcitonin generelated peptide,CGRP)抗体和抗轴突神经丝SMI-312抗体购于Abcam;抗神经元核抗原(neuronal nuclei,NeuN)抗体和抗囊泡γ-氨基丁酸转运体(vesicularγaminobutyric acid transporter,VGAT)抗体购于 Synaptic Systems;抗小胶质细胞抗体(CD11b/c,OX-42)和抗VGLUT2抗体购自Millipore;抗异凝集素B4(isolectin B4,IB4)抗体购于Vector Labs;Alexa Fluor(AF)488、AF546、AF555和AF647标记的荧光Ⅱ抗均购于Molecular Probes;Cy5和rhodamine标记的荧光Ⅱ抗购于Jackson ImmunoResearch Labs;冰冻切片机(CM1950)购自Lecia。激光共聚焦显微镜(LSM700)及图像分析软件(ZEN 2010)购自 Carl Zeiss。
3 主要方法
3.1 免疫组织化学染色脊髓切片制备 给大鼠腹腔注射乌拉坦(1.5 g/kg)麻醉后开胸暴露心脏,用预冷的0.9%NaCl 100 mL行心脏灌流,再以4%多聚甲醛(pH 7.4,4℃)150 mL灌注固定,灌毕立即剪开大鼠背部皮肤,分离脊柱两侧肌肉、离断两侧肋骨,取出连同脊髓的脊柱,并完全浸入4%多聚甲醛中。以腹侧入路切除椎板,以腰骶膨大部为中心游离出脊髓(腰1~骶3),于显微镜下剥离脊髓表面硬脊膜、软脊膜及附着神经根。取出脊髓放入4%多聚甲醛中固定6 h,充分漂洗后置于30%蔗糖溶液3 d(4℃),取出脊髓,切去脊髓头尾,留取L4~L5腰骶膨大部,OCT包埋剂包埋,用冰冻切片机连续横向切片(片厚30 μm),放置于0.01 mol/L磷酸盐缓冲液(phosphatebuffered saline,PBS)液中存放,按下述方法进行免疫荧光染色。
3.2 免疫荧光组织化学染色方法 将上述脊髓横切片在0.01 mol/L PBS液中充分漂洗后进行常规漂染:Ⅰ抗 HCN1~4(1∶200)分别与神经元标志物NeuN(1∶200)、星形胶质细胞标志物 GFAP(1∶2 000)、小胶质细胞标志物OX-42(1∶200)、非肽能初级传入轴突末梢标志物IB4(1∶100)、肽能初级传入轴突末梢标志物CGRP(1∶100)、神经元树突标志物微管相关蛋白2(microtubule-associated protein 2,MAP2;1∶1 000)、神经元轴突标志物 SMI-312(1∶1 000)和抑制性中间神经元轴突末梢标志物VGAT(1∶1 000)进行共染,于4℃摇床上孵育3 d。挑出脊髓片用PBS漂洗3次,每次5 min,加入与Ⅰ抗种属对应的Ⅱ抗 AF488(1∶500)、AF546(1∶400)、AF555(1∶400)、AF647(1∶400)、rhodamine(1∶1 000)和Cy5(1∶400)于4℃摇床上孵育过夜,取出脊髓片充分漂洗后封片。用激光共聚焦显微镜采集10倍(×10)、40倍(×40)及63倍(×63,油镜)的图像进行分析处理,根据实际情况调节 Z值(0.5~1 μm)。HCN1~4抗体的特异性已在我们之前的研究中得到验证[9]。
3.3 荧光定量分析 采用ZEN 2010分析软件自带的荧光定量功能,测定HCN1~4在SDHⅠ~Ⅱ层内侧带、中侧带和外侧带的平均荧光强度(mean fluorescence intensity,MFI),图像均取自同一放大倍数(×10,zoom=1),从每只大鼠切取的脊髓片中随机挑取2~3张,在两侧背角Ⅰ~Ⅱ层内、中、外侧带各取150 pixel×150 pixel正方形方框并测量方框内的MFI,再取同样大小的方框置于空白处以测定背景MFI,用来标准化HCN1~4的MFI(两者相减),取均值作为其MFI。采用上述同种方法,图像取同一放大倍数(×10,zoom=0.5),定量分析HCN1~4在脊髓灰质各层(Ⅰ~Ⅱ层、Ⅲ~Ⅳ层和Ⅴ~Ⅵ层)及白质内的MFI。
3.4 共定位分析 采用ZEN 2010分析软件自带的共染定量分析功能,测定并分析HCN1~4分别与SDH Ⅰ~Ⅱ层各标志物NeuN、GFAP、OX-42、MAP2、SMI-312、CGRP、IB4和VGAT的共定位情况。图像均取自同一放大倍数(×40,zoom=1),从每只大鼠切取的脊髓片中随机挑取2~3张,统计每张脊髓片两侧背角内、中、外侧带400 pixel×400 pixel正方形方框内HCN1~4分别与各标志物的共定位系数,并取其均值作为最终共定位系数。
4 统计学处理
所有计量资料以均数±标准误(mean±SEM)表示。采用SPSS17.0统计软件进行统计学分析,数据经正态性和方差齐性检验后,两组独立样本间的比较采用两独立样本t检验,3组及以上样本间比较采用单因素方差分析。以P<0.05为差异有统计学意义。
结 果
1 HCN通道在脊髓的大体分布
采用免疫组化染色方法,观察HCN1~4在脊髓中的分布模式,结果显示HCN通道的4种亚型均在脊髓灰质中表现出较强的荧光信号,而在白质中荧光信号微弱,见图1A~D。进一步荧光定量分析结果显示HCN1~4在脊髓背角Ⅰ~Ⅱ层内侧带、中侧带和外侧带的MFI无显著差异(P>0.05),见图1E。

Figure 1.Distribution of HCN1~4 in the spinal cord.A~D:representative images of immunolabeling of HCN1,HCN2,HCN3 and HCN4 in the spinal cord(scale bar=200μm);A1~D3:the enlargements of the areas showing in the box in A~D(from top to bottom),respectively(scale bar=20 μm);E:the mean fluorescence intensity(MFI)of HCN1~4 in the lateral,central and medial dorsal horn.Mean±SEM.n=11.图1 HCN通道亚型在脊髓的大体分布
此外,HCN2~4主要集中分布在脊髓背角Ⅰ~Ⅱ层,其MFI与其它各层及白质之间均有显著性差异(P<0.01);而HCN1灰质MFI虽与白质的MFI相比有显著差异(P<0.01),但其Ⅰ~Ⅱ层MFI与其它各层的MFI相比,无显著差异(P>0.05),见表1。

表1 HCN1~4在脊髓背角Ⅰ~Ⅵ层和白质中分布的平均荧光强度Table 1.The MFIof HCN1~4 in laminaeⅠ~Ⅵ and white matter of the spinal cord(Mean±SEM)
2 HCN通道在脊髓背角神经元及胶质细胞上的分布
为明确HCN1~4蛋白表达的细胞类型,我们观察了HCN1~4与NeuN和GFAP、OX-42的共染情况,见图2及表2、3。有11.2%的GFAP、8.7%的OX-42和4.8%的NeuN与HCN1共染;31.7%的NeuN和5.8%的GFAP与HCN2共染,OX-42与HCN2共染少(3%);38.6%的NeuN与HCN3共染,而GFAP和OX-42与HCN3几乎无共染(均<1%);7.2%的GFAP与HCN4共染,NeuN和OX-42与HCN4共染少(分别为1.5%和5%)。此外,NeuN与HCN1~4的共染占HCN1~4的百分比分别为12.6%、45.6%、64.7%和2.2%,GFAP与HCN1~4的共染占HCN1~4的百分比分别为5.2%、2.3%、0.4%和3%,OX-42与HCN1-4的共染占HCN1~4的百分比分别为3.5%、1.3%、0.4%和1.6%。
3 HCN通道在脊髓背角初级传入神经末梢的分布
通过检测HCN通道在脊髓背角初级传入神经末梢的分布观察到IB4与HCN4存在共染,而与HCN1~3几乎不共染,见图3A。CGRP与HCN2和HCN4存在共染,与HCN1和HCN3不共染,见图3B。
定量分析结果表明,IB4和CGRP与HCN通道各亚型共定位存在显著差异(P<0.05),其中6.7%的IB4与HCN4共染,9.2%的CGRP与HCN2共染,IB4、CGRP与HCN1共染少(分别为1.1%和2.1%),而IB4和CGRP与HCN3均无共染(分别为0.3%和0.9%),见表2。此外,9.5%的HCN4与IB4共染,6.3%的HCN2与CGRP共染,见表3。
4 HCN通道在脊髓背角中间神经元树突和轴突的分布
HCN1、HCN4与MAP2存在共染,HCN2、HCN3与MAP2几乎无共染;SMI-312与HCN1~4共染均不显著,见图4。
定量分析结果显示,6%和8.7%的MAP2分别与HCN1和HCN4共染,MAP2与HCN2和HCN3的共染系数分别为2.8%和0.5%;SMI-312与HCN1~4的共染系数分别为3.8%、1.2%、0.5和4.4%,见表2。此外,8.9%的HCN4与MAP2共染,而2%的HCN1与SMI-312共染,见表3。
5 HCN通道在脊髓背角中间神经元轴突末梢的分布
为明确HCN1~4在抑制性轴突末梢的分布情况,我们将其与抑制性中间神经元轴突末梢标志物VGAT共染,见图5。
定量分析结果显示,15.9%的VGAT与HCN4共染,而VGAT与HCN1~3几乎不共染(分别为2.3%、1.6%和0.5%);同时14.7%的HCN4共表达VGAT,见表2、3。
讨 论
1 HCN通道在脊髓的大体分布
背角浅层是伤害性信息由外周传入高级中枢的第一个中继站,Ⅰ层由投射神经元和中间神经元组成,Ⅱ层的SG区则是“闸门控制理论”的核心组成部分,闸门的开放与关闭决定了疼痛信息能否继续向上传递,在慢性疼痛的发生与发展过程中具有至关重要的作用[10-12]。因此,HCN通道在脊髓背角浅层的表达尤为被学者们关注。在成年雄性Wistar大鼠或C57BL/6小鼠中,HCN2主要位于脊髓背角的Ⅰ和Ⅱ外层(Ⅱo)[13-14],HCN4 则主要位于Ⅰ和Ⅱ内层(Ⅱi)[15]。与上述结果一致,我们近年已发表的工作表明HCN亚型在脊髓背角Ⅰ~Ⅵ各层均有分布,且以Ⅰ~Ⅱ层为主[9]。由于SG神经元随着放电模式的不同,在脊髓背角呈现不同的分布模式,如适应性放电神经元主要分布在SG区的外侧带[16]。因此,有必要明确HCN1~4在脊髓背角浅层的内、中、外侧带是否存在差异性表达。本研究结果显示它们之间的分布无显著差异,进一步的定量分析显示,HCN2~4在脊髓背角的分布主要以Ⅰ~Ⅱ层为主。由于Ⅰ~Ⅱ层与痛信号的传递密切相关,因此HCN通道在该区域的高表达表明其在疼痛的调制中可能具有重要的作用。

Figure 2.Specific localization of HCN isoforms in different cell types in spinal dorsal horn.A:representative images of co-immunostaining for HCN1~4(green)and NeuN(red)in superficial spinal dorsal horn;B:representative images of co-immunostaining for HCN1~4(green),GFAP(blue)and OX-42(red)in superficial spinal dorsal horn.图2 HCN亚型在脊髓背角细胞中的特异性表达
2 HCN通道在脊髓背角细胞水平上的分布

Figure 3.Distribution of HCN isoforms in the primary afferent nerve endings of the spinal dorsal horn.Representative images of coimmunostaining for HCN1~4(green),IB4(blue)and CGRP(red)in superficial spinal dorsal horn were shown.图3 HCN通道各亚型在脊髓背角初级传入神经末梢的分布
脊髓背角主要由神经元和胶质细胞组成,后者主要包括星形胶质细胞和小胶质细胞。活化的胶质细胞通过释放多种炎性细胞因子如肿瘤坏死因子α和白细胞介素1β等致神经病理性疼痛胶质细胞的激活是神经痛发病的重要因素,葛根素[17]及小胶质细胞抑制剂米诺环素[18]等可减少这些致痛物质的生成以有效缓解疼痛。我们已发表的研究观察到米诺环素可在数分钟内显著降低SG神经元的HCN通道介导的电流(Ih)的振幅从而降低其兴奋性[19],证实了米诺环素镇痛的神经机制。那么米诺环素有无可能通过作用于胶质细胞的HCN通道而发挥作用呢?在大鼠的大脑皮质和海马中,Honsa等[20]检测到这两个区域的星形胶质细胞在生理情况下表达极少量的HCN1~3,在缺血刺激下,这些细胞的HCN1~3可显著增加,而HCN4在生理和病理情况下均未发现有表达。但是,HCN通道在脊髓背角的胶质细胞是否存在表达尚不明确。本研究首次揭示正常情况下脊髓背角的星形胶质细胞表达HCN1、HCN2和HCN4通道,而HCN3与星形胶质细胞无共定位。由于病理性疼痛状态下星形胶质细胞显著激活,HCN通道的表达有可能如上述缺血刺激亦大幅上升,从而加重疼痛。另一方面,虽然脊髓背角HCN1与小胶质细胞标志物OX-42存在少量共染,但其余3种亚型与OX-42均几乎无共定位,表明小胶质细胞的HCN通道对疼痛的作用有限。此外,多个电生理研究均已在背角浅层神经元上记录到Ⅰh[19,21],本研究观察到NeuN共染较多的亚型为HCN2和HCN3。研究表明,脊髓背角浅层的神经元多为中间神经元[22],84.1%小清蛋白阳性神经元和53.9%的PKCγ阳性神经元被检测有HCN4表达[15],此外,HCN4还被检测到表达在不含钙结合蛋白的Pax2阳性的背角Ⅱ层神经元中[23]。目前HCN通道在SDH中间神经元的定位尚不完全清楚,进一步研究HCN通道在脊髓背角中间神经元亚群中的分布,对阐明HCN通道在神经病理性疼痛和炎性痛中的作用具有至关重要的作用。

Figure 4.Distribution of HCN isoforms in dendrites and axons of interneurons in spinal dorsal horn.A:representative images of coimmunostaining for HCN1~4(green)and MAP2(red)in superficial spinal dorsal horn;B:representative images of co-immunostaining for HCN1~4(green)and SMI-312(red)in superficial spinal dorsal horn.图4 HCN通道亚型在脊髓背角神经元树突和轴突中的分布
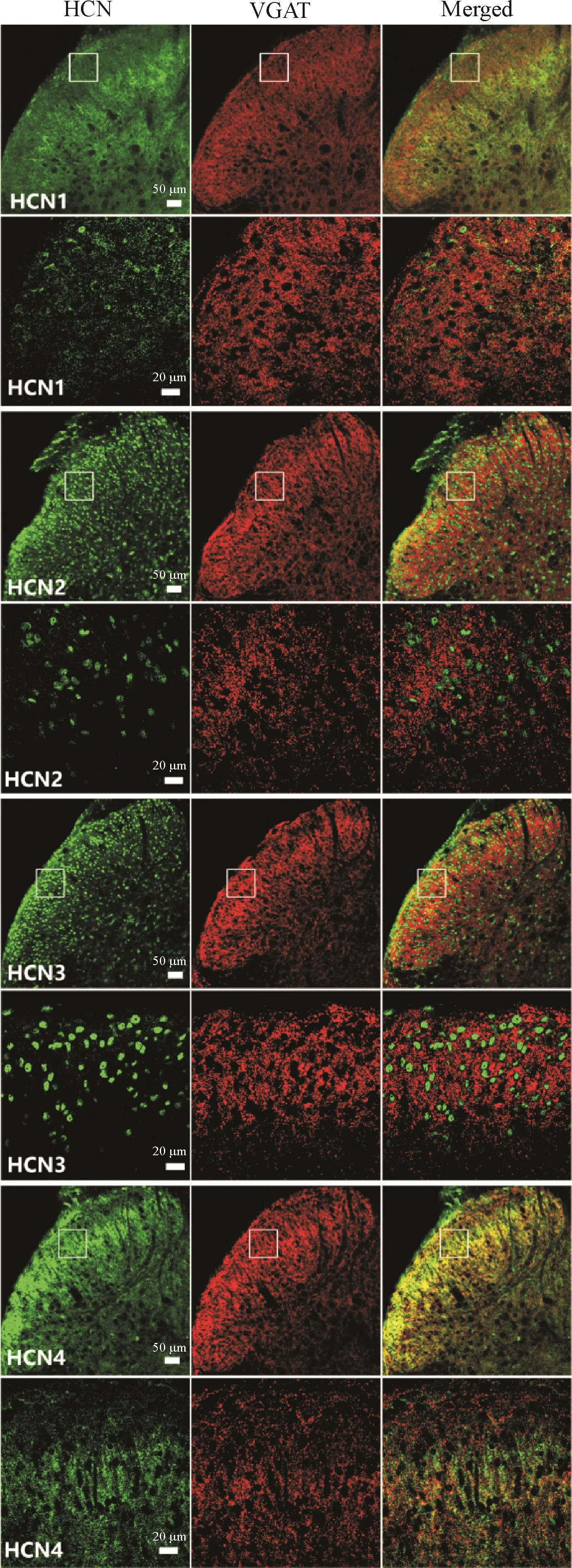
Figure 5.Distribution of HCN isoforms at axonal terminal of inhibitory interneurons in spinal dorsal horn.Representative images of co-immunostaining for HCN1~4(green)and VGAT(red)in superficial spinal dorsal horn were shown.图5 HCN通道亚型在脊髓背角抑制性中间神经元轴突末梢的分布
3 HCN通道在脊髓背角神经元亚细胞水平上的分布
脊髓背角接收来自初级传入轴突的输入信号,其递质主要有肽能(P物质和CGRP)和非肽能(IB4)两种。IB4阳性的传入神经纤维不参与正常的机械伤害感受,但能被炎症刺激致敏并在介导机械性炎症超敏反应中起关键作用[24]。而CGRP标记阳性的肽能初级传入神经元也与炎症性疼痛相关[3]。以往研究表明,HCN2主要与CGRP和SP共染,而与IB4几乎无共染[13-14,25]。与该结果类似,我们亦观察到HCN2主要与CGRP共染,与IB4无共染,同时我们首次发现HCN4与IB4存在共定位,揭示了HCN通道各亚型在伤害性初级传入神经末梢中有其独特的表达模式,其中HCN2和HCN4是分布在伤害性初级传入末梢的主要类型。本课题组在近年的一项研究中已证实HCN1~4亚型在兴奋性突触末端有表达,且以HCN4为主[8]。本研究进一步发现HCN1~4与VGAT均有共定位,HCN4的共定位明显高于其他亚型。以上结果提示,在结构功能和表达上,HCN4与中间神经元轴突末梢存在紧密且复杂的联系。树突和轴突是神经元接收和传出信息的两个主要结构,Jiang等[26]检测到慢性压迫模型诱导的神经病理性痛模型大鼠中,受损坐骨神经的HCN1和HCN2显著增加,表明周围神经的轴突上存在HCN表达。本研究首次发现背角浅层的HCN1和HCN4与树突标志物少量共染,与轴突标志物几乎不共染,可能与背角Ⅱ层的解剖构成有关,即脊髓薄片的Ⅱ层在光学显微镜下表现为半透明状,主要原因是此层的轴突较少。在SMI-312表达稍多的Ⅲ层,HCN1和HCN4均与其存在不同程度的共染。以上结果表明HCN1和HCN4在背角浅层神经元接收信息的过程中发挥一定的作用。
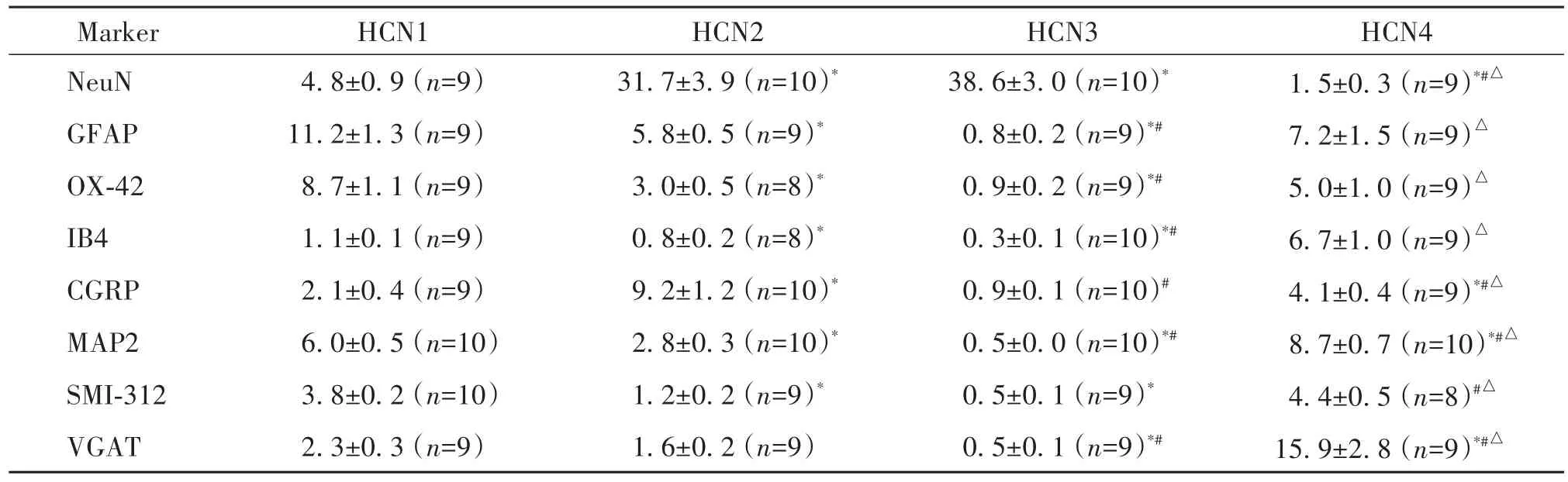
表2 HCN1~4与免疫标志物共染部分占免疫标志物的百分比Table 2.Percentage of marker-IRprofiles immunoreactive for HCN1~4(%.Mean±SEM)

表3 HCN1~4与免疫标志物共染部分占HCN1~4的百分比Table 3.Percentage of HCN1~4-IRprofiles immunoreactive for the markers(%.Mean±SEM)
综上所述,本研究揭示了HCN通道各亚型在大鼠脊髓背角细胞和亚细胞结构上的定位特点,为阐明HCN通道在脊髓水平的生理和病理作用提供了参考资料,有助于深入研究HCN通道在脊髓背角的调控机制。
猜你喜欢
中国疼痛医学杂志(2022年9期)2022-10-08
实用医学杂志(2021年22期)2021-12-28
浙江临床医学(2021年12期)2021-11-29
皮肤病与性病(2021年3期)2021-07-30
中国动物检疫(2020年1期)2020-01-08
中国疼痛医学杂志(2019年9期)2019-09-20
中国兽药杂志(2019年4期)2019-05-15
组织工程与重建外科杂志(2017年1期)2017-01-10
中国疼痛医学杂志(2017年2期)2017-01-10
