
GnIH通过p38MAPK信号通路对猪卵巢颗粒细胞自噬与凋亡的影响
2020-06-03 08:02张鑫霍孔林宋星星张多妮胡文胡传活李珣
中国农业科学 2020年9期
张鑫,霍孔林,宋星星,张多妮,胡文,胡传活,李珣
GnIH通过p38MAPK信号通路对猪卵巢颗粒细胞自噬与凋亡的影响
张鑫,霍孔林,宋星星,张多妮,胡文,胡传活,李珣
(广西大学动物科学技术学院,南宁 530004)
【】细胞自噬和凋亡存在着相互制约,p38MAPK信号通路作为细胞凋亡的主要调控通路之一,也对细胞自噬存在促进和抑制的双重作用。已有研究表明,促性腺激素抑制激素(gonadotropin-inhibitory hormone , GnIH)对细胞自噬与凋亡均有影响,但作用机制尚不明确。故探究GnIH通过p38MAPK信号通路对猪卵巢颗粒细胞(pGCs)自噬与凋亡的影响及其机理,为解决母猪的产子率以及同期发情等问题提供参考。从猪卵巢中提取卵巢颗粒细胞,进行体外培养。1、探究GnIH对p38MAPK信号通路的最佳作用时间:按孵育GnIH时间梯度(0 min、10 min、30 min、60 min、90 min)分组,用Western blot检测猪卵巢颗粒细胞p38与p-p38的蛋白表达量变化;2、验证GnIH对p38MAPK信号通路的影响:按(空白对照、GnIH、p38激活剂(U-46619)、U-46619+GnIH)分组,用Western blot检测p38与p-p38的蛋白表达量变化;3、探究不同浓度GnIH对自噬和凋亡的影响:按(空白对照、10-6mol·L-1GnIH、10-8mol·L-1GnIH、10-10mol·L-1GnIH、10-12mol·L-1GnIH)分组,用Western blot检测自噬与凋亡标志性蛋白的表达量变化;4、验证不同浓度GnIH通过p38信号通路对自噬和凋亡的影响:将细胞分成6组(空白对照、U-46619、U-46619+10-6mol·L-1GnIH、U-46619+10-8mol·L-1GnIH、U-46619+10-10mol·L-1GnIH、U-46619+10-12mol·L-1GnIH),用Western blot检测自噬与凋亡标志性蛋白的表达量变化。1. GnIH孵育10 min后,显著降低p38与p-p38的蛋白表达量(<0.05),提示,GnIH对p38MAPK信号通路的最佳作用时间为10 min;2. U-46619显著促进pGCs的p38磷酸化水平(<0.05),GnIH显著抑制pGCs的p38磷酸化水平(<0.05),提示,U-46619使p38MAPK信号通路活化,GnIH对p38MAPK信号通路的活化有抑制作用;3. 当 GnIH的浓度为10-6mol·L-1时,pGCs的自噬和凋亡水平显著升高(<0.05),随着GnIH浓度的降低,pGCs的自噬水平逐渐升高(<0.05),pGCs的凋亡水平逐渐降低(<0.05),提示,高浓度GnIH可以促进自噬和凋亡,随着GnIH浓度的降低,自噬水平逐渐升高,而凋亡水平逐渐下降;4. 加入U-46619后,GnIH使pGCs的自噬显著上调(<0.05),并且使pGCs的凋亡显著下调(<0.05),提示,不同浓度GnIH通过p38MAPK信号通路影响pGCs的自噬和凋亡。GnIH可能通过抑制p38MAPK信号通路的活化,上调pGCs的自噬,减少pGCs的凋亡。
自噬;凋亡;GnIH;p38MAPK;猪;卵巢颗粒细胞
0 引言
【研究意义】卵巢闭锁是哺乳动物卵巢中普遍存在的现象[1]。卵巢闭锁对母猪的产子率有影响。促性腺激素抑制激素(gonadotropin-inhibitory hormone,GnIH)作为一种由下丘脑释放的性腺激素,我们猜测,对卵巢细胞活性有着一定的调控作用。为此,本试验可以为解决母猪的产子率以及同期发情[2]等问题提供参考。【前人研究进展】GnIH是Tsutsui等于2000年从鹌鹑的下丘脑中分离的一种C末端具有RF酰胺结构的12肽[3]。后续研究发现哺乳动物中存在GnIH类似物-RF酰胺相关肽(RFRP),RFRP 基因主要编码两条有生物活性的短肽,分别为RFRP-1、RFRP-3,然而在大多数哺乳动物生殖系统中发挥主要作用的是RFRP-3[4],也是目前研究较为广泛的[5-7]。人类和其他物种的研究数据表明,RFRP-3可能参与病理变化,是影响排卵、卵泡发育和卵巢分泌类固醇的因素[8-9]。GnIH可以通过对下丘脑GnRH和垂体前叶的作用抑制促性腺激素释放[10]。GnIH在生殖调控中具有重要作用,在其他脊椎动物中也具有类似的作用[11]。细胞的活性决定各项生理机能,由此凋亡作为第一个细胞程序性死亡被认知[12],并成为哺乳动物组织细胞死亡的主要机制。但近来研究发现凋亡并不能单独对细胞死亡起决定性作用,由此自噬被发现[13],并探究出自噬与凋亡存在促进与抑制的双重作用。已有研究证明,肿瘤细胞中自噬可以使凋亡率下降[14]。自噬与凋亡也可以共同导致细胞死亡,当自噬或凋亡受到抑制,另外一种则会转变成细胞的死亡途径[15-16]。当自噬体积累到一定水平,则会对细胞的凋亡产生影响。卵母细胞质量是由卵泡闭锁这一正常的生理过程决定的,而细胞的自噬与凋亡可能参与调节卵泡闭锁[17]。研究表明,卵泡的发育和颗粒细胞的凋亡过程中都存在着细胞自噬[18],因此自噬可能参与颗粒细胞的凋亡;大鼠颗粒细胞的自噬可能与卵泡发育有关[19],Choi在研究自噬与卵泡闭锁的关系中发现,在卵泡的发育过程中,自噬主要集中在卵巢颗粒细胞,并且与细胞的凋亡存在着相关性[20]。在卵泡闭锁的初期,颗粒细胞中有Beclin-1的表达,而Caspase-3则是分布在卵泡中;在中期,卵巢颗粒细胞中Beclin-1和Caspase-3均有表达;而在晚期,颗粒细胞中Caspase-3的表达水平较高[19]。LC3在大鼠卵泡颗粒细胞中表达量最高, 也表明自噬主要发生在颗粒细胞中, 而自噬导致的颗粒细胞死亡可能参与卵泡闭锁[21]。p38MAPK信号通路是细胞调节的主要信号通路之一[22],MAPK信号通路可能参与调节GCs的凋亡[23]。GnIH参与调节小鼠卵巢的发育及功能[24-25]。目前已证实,GnIH对母猪生殖有一定的调控作用[26]。MADDINENI于2008年发现GnIH能减少鸡卵巢颗粒细胞的活化,但是这种抑制作用能被外源FSH中和[27-28]。在睾丸细胞中,GnIH与GnIH-R均有表达[29],表明GnIH可能参与生殖细胞的分化。给成熟鹌鹑连续14 d注射GnIH,发现睾丸中精原细胞活性降低,并诱导细胞凋亡[30],提示GnIH可能通过诱导细胞凋亡从而调节性腺的发育。研究表明,GnIH对小鼠附睾组织细胞的凋亡和自噬有影响[31]。【本研究切入点】虽然目前已有关于GnIH在卵巢水平的相关研究。但是GnIH处理的pGCs通过p38MAPK信号通路对细胞凋亡与自噬的调控还未见报道。因此,我们提出猜测,GnIH可能通过p38MAPK信号通路对细胞凋亡与自噬产生影响。【拟解决的关键问题】首先探究了GnIH是否对p38MAPK信号通路的活化产生影响;进一步探究了GnIH是否对卵巢颗粒细胞的自噬和凋亡有影响;最后探究了GnIH是否是通过p38MAPK信号通路对自噬和凋亡产生的影响。
1 材料与方法
1.1 材料和试剂
本试验于2018年3—10月在广西大学解剖组培实验室完成。
DMEM/F12培养基、澳洲来源胎牛血清购自南京维森特生物技术有限公司。台盼蓝染液购自Gibco。其他试剂均为国产分析纯。Western细胞裂解液、超敏ECL化学发光试剂盒、BCA蛋白定量试剂盒购于上海碧云天生物技术有限公司,一抗GAPDH、p38、p-p38、Bcl-2、Caspase3、Bax、Beclin-1、LC3、Atg5和Atg12浓度均为1﹕2 000,二抗羊抗鼠、羊抗兔浓度为1﹕20 000均购自CST公司。GnIH由实验室于NCBI查找相关氨基酸序列,送至公司合成。
1.2 猪卵巢的采集
从广西本地屠宰场当天取回卵巢,将卵巢放置在含有两性霉素(青霉素/链霉素(1%))的生理盐水中(37℃),2 h内运输回广西大学解剖组培实验室。用生理盐水冲洗备用。
1.3 猪卵巢颗粒细胞的体外培养
pGCs分离培养方法参考相关研究[32-33]进行。于37℃、5%CO2及饱和湿度的培养箱中进行成熟培养,培养48 h后,加入p38MAPK激活剂U-46619和不同浓度的GnIH(购自康肽生物(北京)科技有限公司),培养1 h后,收取细胞,每组试验平行3次重复。
1.4 猪卵巢颗粒细胞蛋白的提取
弃去细胞瓶中培养液,加入PBS,洗2—3次,弃洗液;加入混有1%PMSF的细胞裂解液,静置30 min,每隔10 min轻轻晃动细胞瓶;裂解后,将细胞瓶内裂解液移入1.5 mL预冷的EP管,4℃、12 000 r/min离心5 min;BCA测浓度,分装,-80℃保存。
1.5 Western blot检测
12%SDS-PAGE凝胶电泳,半干法转印至PVDF膜上,放入5%脱脂牛奶,室温封闭1—2 h,一抗p38、p-p38、Bcl-2、Caspase3、Bax、Beclin-1、LC3、Atg5和Atg12以及GAPDH,4℃过夜,二抗孵育1 h,每步完成后,PBST洗涤3次,每次15 min。ECL化学发光液显色,暗室曝光,扫描灰度值。试验重复3次,用 ImageJ进行灰度值分析,其中GAPDH作为标准化的内部对照。
1.6 数据分析
所有数据均显示为平均数±标准误(Mean±SE)。使用SPSS 21.0统计软件进行单因素方差分析,LSD检验各组之间的统计学显著性。检验结果以*<0.05为差异显著,**<0.01为差异极显著。
2 结果
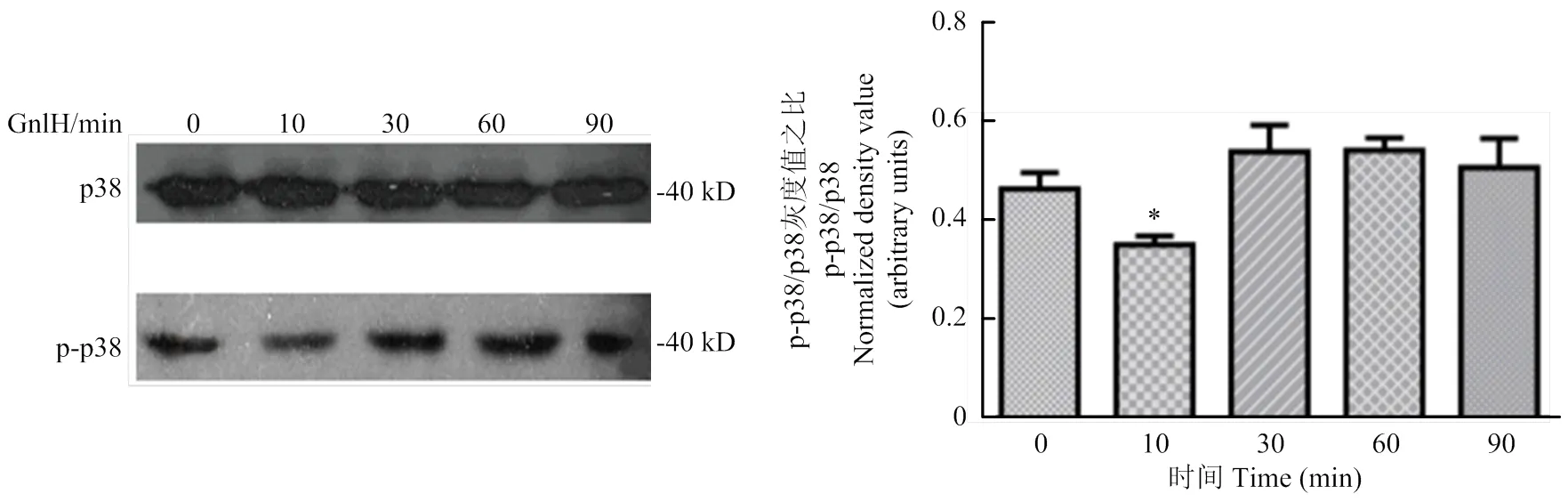
2.1 GnIH对猪卵巢颗粒细胞p38MAPK信号通路的最佳作用时间
在GnIH梯度时间(0、10、30、60、90 min)孵育pGCs后,采用Western blot检测p38及p-p38的表达变化,检测灰度值。结果提示10 min与其他试验组比较,p38及p-p38显著降低(<0.05)(图1)。
2.2 GnIH对p38MAPK信号通路的影响
Western blot检测(空白、GnIH、p38激活剂、p38激活剂+GnIH)p38及p-p38的表达变化,检测灰度值。GnIH组与空白组比较,p38及p-p38的比值显著降低(<0.05);p38激活剂组与空白组比较,p38及p-p38的比值显著升高(<0.05);p38激活剂+GnIH组与p38激活剂比较,p38及p-p38的比值显著降低(<0.05,图2)。
2.3 不同浓度GnIH对猪卵巢颗粒细胞自噬和凋亡的影响
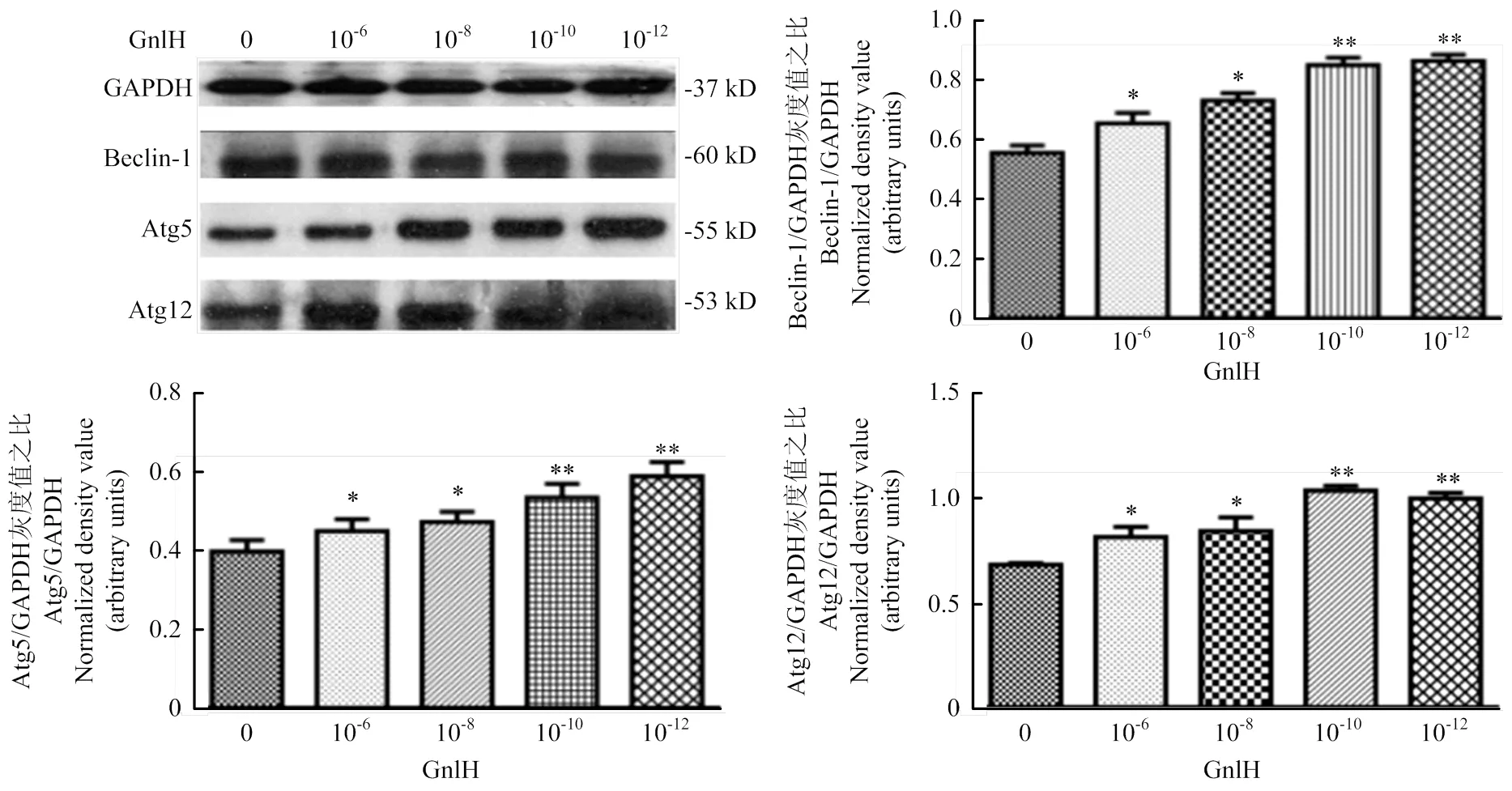
GnIH浓度梯度(0、10-6、10-8、10-10、10-12)处理pGCs,Western blot检测Caspase-3、Bax和Bcl-2,灰度值分析。caspase-3、Bcl-2、Bax和GAPDH的免疫印迹在大约32、26、21和37 kD。试验组与对照组相比均有显著性差异(<0.05),结果提示,低剂量组Bcl-2的表达极显著升高(<0.01),caspase-3与Bax的表达极显著降低(<0.01,图3)。
Western blot测定Beclin-1、Atg12和Atg5,灰度值分析。Beclin-1、Atg5和Atg12的免疫印迹分别位于60、55和53 kD。结果提示,低剂量组Beclin-1、Atg12和Atg5的表达极显著升高(<0.01,图4)。
2.4 不同浓度GnIH通过p38MAPK信号通路对猪卵巢颗粒细胞自噬和凋亡的影响
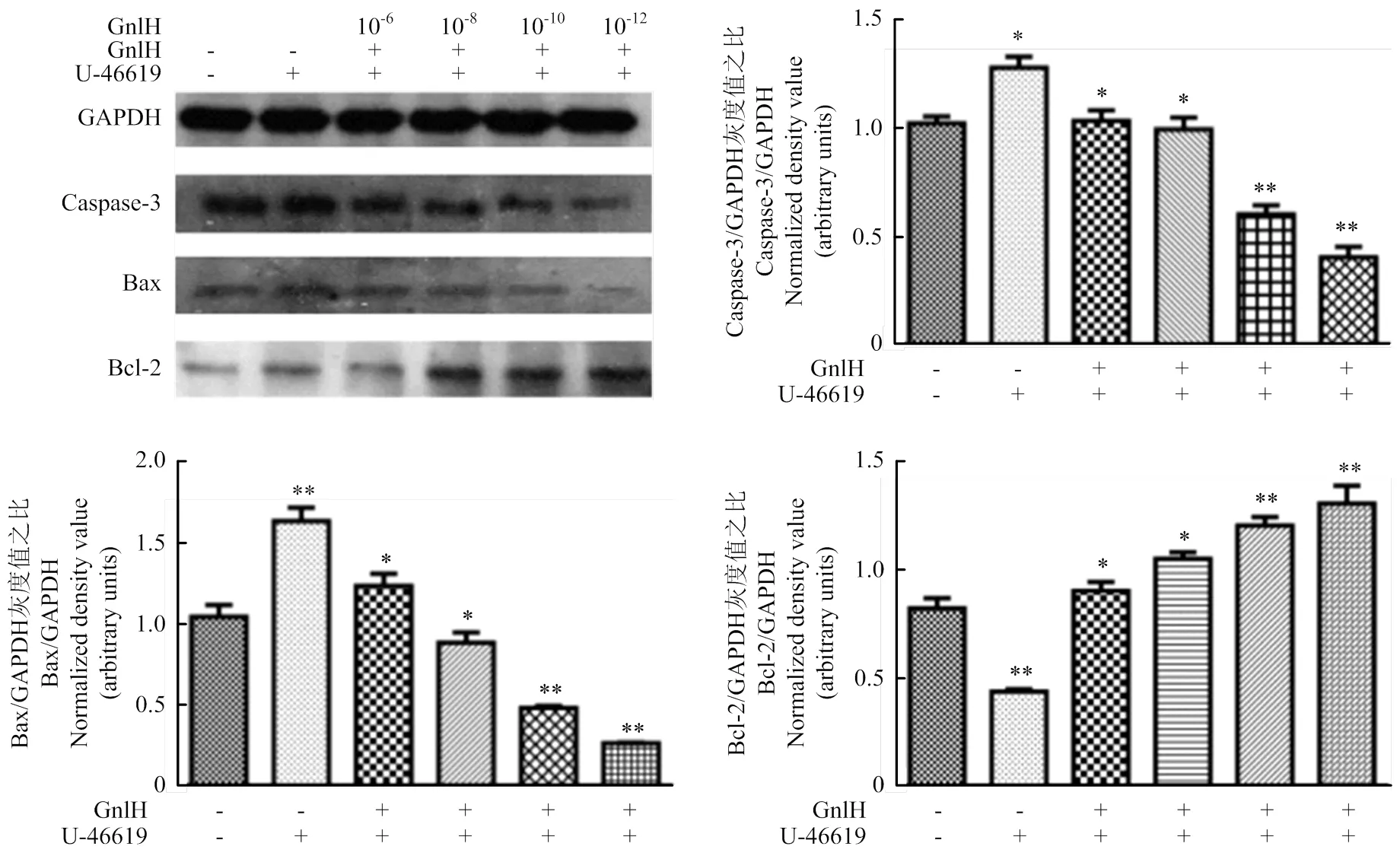
p38激活剂孵育pGCs后,GnIH浓度梯度(0、10-6、10-8、10-10、10-12)处理pGCs,Western blot检测Caspase-3、Bax和Bcl-2,灰度值分析。结果提示,P38激活剂组与空白对照相比,存在显著性差异(<0.05);经不同剂量GnIH处理后的试验组与p38激活剂组比,低剂量组Bcl-2的表达极显著升高(<0.01);caspase-3与Bax的表达极显著降低(<0.01,图5)。
用ImageJ进行灰度值分析。数值为平均值±S.E,与对照组比较,星号表示显著差异。*P<0.05;**P<0.01

用ImageJ进行灰度值分析。数值为平均值±S.E,与对照组比较,星号表示显著差异。*P<0.05;**P<0.01

用ImageJ进行灰度值分析。数值为平均值±S.E,与对照组比较,星号表示显著差异。*P<0.05;**P<0.01
用ImageJ进行灰度值分析。数值为平均值±S.E,与对照组比较,星号表示显著差异。*P<0.05;**P<0.01
用ImageJ进行灰度值分析。数值为平均值±S.E,与对照组比较,星号表示显著差异。*P<0.05;**P<0.01
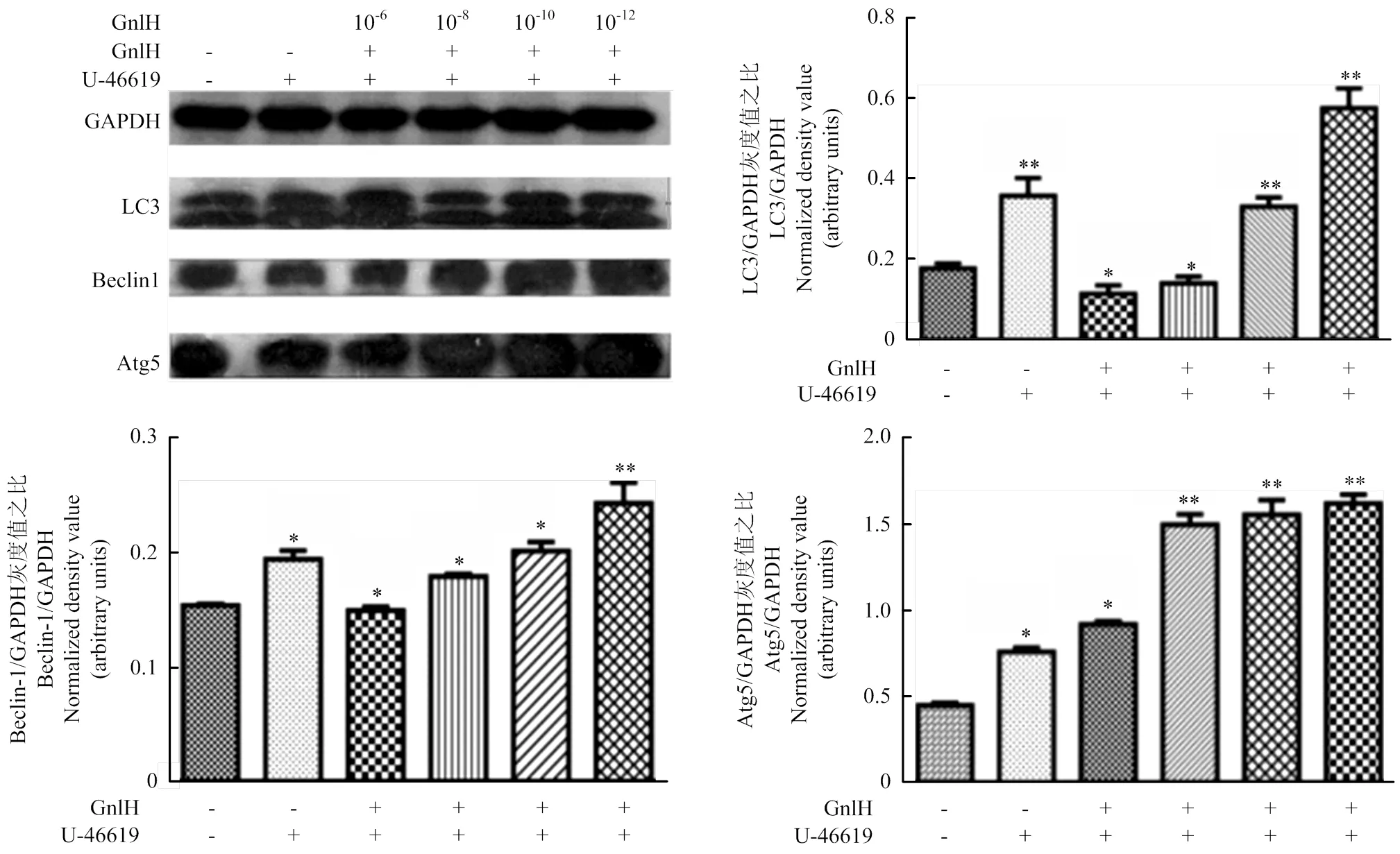
Western blotting测定Beclin-1、LC3和Atg5,灰度值分析。结果显示,P38激活剂组与空白比较,存在显著性差异(<0.05);经不同剂量GnIH处理后的试验组与p38激活剂组比较,均有显著性差异(<0.05);低剂量组Beclin-1、LC3和Atg5的表达极显著升高(<0.01,图6)。
3 讨论
相关研究表明,在GT1-7 细胞系上,mRFRP3预处理可使vip诱导的p38磷酸化受到抑制,此结果显示GnIH具有抑制p38磷酸化的作用[34]。本研究中,体外培养pGCs,加入GnIH孵育,检测其对p38MAPK信号通路活化的影响。当GnIH作用10 min时,对p38MAPK信号通路活化的影响最显著(<0.05);加入p38激活剂后,GnIH可以有效抑制p38MAPK信号通路活化。与前人的结论一致,GnIH可以抑制p38磷酸化。
已有研究表明,在卵巢颗粒细胞中存在着细胞自噬与凋亡。我们猜想GnIH可能与颗粒细胞的自噬和凋亡有关。结果提示,高浓度GnIH可以促进细胞的自噬和凋亡,随着GnIH浓度的降低,凋亡水平逐渐降低,而自噬水平逐渐升高。2017年Dave等对动情前期小鼠卵巢进行了研究,研究发现,RFRP-1使Caspase-3、凋亡标记物明显减少[35]。与本试验结果一致。前期实验室已证明GnIH可以调控附睾的自噬与凋亡[33],GnIH对附睾凋亡出现了极显著的促进作用,而对自噬则出现了双向调控。与本试验结果不一致。猜测可能原因卵巢颗粒细胞是自噬的主要场所,有研究证明,细胞可以通过促进自噬来抑制凋亡的产生,从而提高细胞活性;附睾为体内试验,猜测动物体内对GnIH有神经激素调节作用,导致结果不同;通过试验验证,激素的浓度与作用时间均会对细胞的自噬与凋亡产生影响。
p38MAPK磷酸化水平升高,发挥凋亡促进的作用[36-37],研究证实,抑制p38MAPK信号通路可以通过阻止COCs扩散,从而影响GCs的生长,该结论在大鼠GCs和卵母细胞选择性敲除p38受体中也得到了证实[38];INAGAKI等发现大鼠GCs中存在p38受体,用p38抑制剂SB203580抑制大鼠GCs的p38受体,试验显示通过降低FSH,从而降低了雌二醇和孕激素的产生,充分证明p38及其受体在GCs中发挥重要作用,进而促进卵母细胞成熟。当p38MAPK信号通路被激活,能通过调节细胞形态和提高细胞存活率促进细胞分化[39]。本试验加入p38MAPK激活剂,结果提示,p38MAPK激活剂可以促进p38MAPK的磷酸化;p38MAPK激活剂可以促进卵巢颗粒细胞的凋亡和自噬,与前人研究一致。加入GnIH后,结果提示,不同浓度的GnIH对p38MAPK信号通路活化都具有抑制作用。我们进一步探究了p38MAPK信号通路活化后,GnIH对卵巢颗粒细胞自噬和凋亡的影响,从而探究GnIH是否通过p38MAPK信号通路对卵巢颗粒细胞的自噬和凋亡进行调控。通过检测自噬和凋亡的标志性蛋白,我们得出,随着GnIH浓度的降低,Caspase-3和Bax的表达呈下降趋势,Bcl-2的表达呈上调趋势,提示了GnIH通过抑制p38MAPK信号通路活化对pGCs的凋亡存在高浓度促进,低浓度抑制的作用;Beclin-1、Atg12和Atg5的表达呈上调趋势,提示了GnIH对通过抑制p38MAPK信号通路活化对自噬有促进作用,GnIH的浓度越低,促进作用越明显,且浓度低于10-10mol·L-1,有极显著的促进作用(<0.01)。以往研究表明,自噬的作用不仅通过增强caspase的活化导致细胞凋亡,还可以通过凋亡导致细胞死亡。通过应激反应,延缓caspase的激活,可以清除受损细胞[40],这些数据证明了细胞凋亡和自噬之间存在着明显的复杂性,有时甚至相互矛盾。
数值为平均值±S.E,与对照组比较,星号表示显著差异。*P<0.05;**P<0.01
4 结论
本研究证明了促性腺激素抑制激素可以通过调控p38MAPK信号通路影响猪卵巢颗粒细胞的凋亡和自噬。自噬和凋亡的关系异常复杂,自噬和凋亡又存在很多交叉的信号通路。因此,本试验的结果为后续研究猪卵巢颗粒细胞自噬与凋亡的分子机制提供了参考,也为研究卵泡闭锁提供了新的思路。
[1] 谢敏, 姜法贵, 郭燕君. 卵泡发育和闭锁过程中颗粒细胞自噬和凋亡关系的研究. 四川生理科学杂志, 2015, 37(2): 85-88.
XIE M, JIANG F G, GUO Y J. Study on the relationship between autophagy and apoptosis of granulosa cells during follicular development and atresia.2015, 37(2): 85-88. (in Chinese)
[2] 赵红秋. 浅析提高母猪生产性能的措施. 今日畜牧兽医, 2019, 35(4):69.
ZHAO H Q. Analysis on the measures to improve the performance of sows., 2019, 35(4): 69. (in Chinese)
[3] Tsutsui K, Saigoh E, Ukena K, Teranishi H, Fujisawa Y, Kikuchi M, Ishii S, Peter J. A novel avian hypoth- alamic peptide inhibiting gonadotropin release., 2000, 275(2): 661-667.
[4] Tsutsui K, Ubuka T, Bentley G E, Kriegsfeld L . Gonadotropin-inhi-bitory hormone(GnIH): discovery, progress and prospect., 2012, 177(3): 305-314.
[5] 龚金秋, 阳美霞, 唐娇美, 曾杰, 王水莲. RFRP-3对哺乳动物生殖激素的调节作用. 中兽医医药杂志, 2017(4):31-33.
GONG J Q, YANG M X, TANG J M, ZENG J, WANG S L. Regulatory effect of RFRP-3 on reproductive hormones in mammals.2017 (4): 31-33. (in Chinese)
[6] 韩兴绘, 俞建. RFRP-3/GPR147信号通路在哺乳动物生殖系统中作用的研究进展. 中国中西医结合儿科学, 2016, 8(3): 253-257.
HAN X H, YU J. Progress in the study of the role of RFRP-3/GPR147 signaling pathway in the reproductive system of mammals.2016, 8 (3): 253-257. (in Chinese)
[7] Kim J S, Brownjohn P W, Dyer B S, Beltramo M, Walker C S, Hay D L, Painter G F, Tyndall J D, Anderson G M. Anxiogenic and stressor effects of the hypothalamic neuropeptide rfrp-3 are overcome by the NPFFR antagonist GJ14., 2015, 156(11): 4152-4162.
[8] Bentley G E, Ubuka T, McGuire N L, Calisi R, Perfito N, Kriegsfeld L J, Wingfield J C, Tsutsui K. Gonadotrophin- inhibitory hormone: a multifunctional neuropeptide., 2009, 21 276-281.
[9] Bentley G E, Wilsterman K, Ernst D K, Lynn S E, Dickens M J, Calisi R M, Kriegsfeld L J, Kaufer D, Geraghty A C, McGuire N L, Lopes P C, Tsutsui K. Neural versus gonadal GnIH: are they independent systems A mini-review., 2017, 57 1194-1203.
[10] Rizwan M Z, Harbid A A, Inglis M A, Quennell J H , Anderson G M. Evidence that hypothalamic RFamide related peptide-3 neurones are not leptin-responsive in mice and rats., 2014, 26: 247-257.
[11] Li X, Su J, Lei Z, Zhao Y Y, Jin M M, Fang R, Zheng L C, Jiao Y. Gonadotropin-inhibitory hormone (GnIH) and its receptor in the female pig: cDNA cloning, expression in tissues and expression pattern in the reproductive axis during the estrous cycle., 2012, 36(2): 176-185.
[12] Kerr J F, Wyllie A H, Currie A R. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics1972, 26(4): 239-257.
[13] Obeid L M , Linardic C M , Karolak L A , & Hannun Y A . Programmed cell death.1993, 259(5102), 1769-1771.
[14] Zhang H, Kong X X, Kang J S, Su J, Li Y, Zhong J T, Sun L K. Oxidative stress induces parallel autophagy and mitochondria dysfunction in Human Glioma U251 Cells., 2009. 110(2): 376-388.
[15] 汪宇, 周桃, 孙韩艳, 黄蓓, 姜黄素类似物EF24诱导A549细胞自噬及凋亡关系的研究. 中国细胞生物学学报, 2012, 34(6): 590-596.
WANG Y, ZHOU T, SUN H Y, HUANG P. Study on the relationship between autophagy and apoptosis in A549 cells induced by curcumin analogue EF24.2012, 34(6): 590-596. (in Chinese)
[16] Eisenberg-Lerner A, Bialik S, Simon H U, Kimchi A. Life and death partners: apoptosis, autophagy and the cross-talk between them., 2009, 16(7): 966-975.
[17] Choi J, Jo M, Lee E, Choi D. Induction of apoptotic cell death via accumulation of autophagosomes in rat granulosa cells.2011, 95(4): 1482-1486.
[18] Cardenas H, Pope W F. Control of ovulation rate in swine., 2002, 80: 36-46.
[19] Choi J Y, Jo M W, Lee E Y, Yoon B K, Choi D S. The role of autophagy in follicular development and atresia in rat granulosa cells.2010, 93(8): 2532-2537.
[20] Hułas-Stasiak M. Gawron A. Follicular atresia in the prepubertal spiny mouse () ovary., 2011, 16(10): 967-975.
[21] Morais R D V S, Thomé R G, Lemos F S, Rizzo N B E. Autophagy and apoptosis interplay during follicular atresia in fish ovary: a morphological and immunocytochemical study., 2012, 347(2): 467-478.
[22] Tamura M, Nakagawa Y, Shimizu H, Yamada N, Miyano T, Miyazaki H. Cellular functions of mitogen-activated protein kinases and protein tyrosine phosphatases in ovarian granulosa cells., 2004 Feb; 50(1): 47-55.
[23] Villadiaz L G, Miyano T. Activation of p38 MAPK during porcine oocyte maturation., 2004, 71(2): 691-696.
[24] Singh P, Krishna A, Tsutsui K. Effects of gonadotropin- inhibitory hormone on folliculogenesis and steroidogenesis of cyclic mice., 2011, 95(4): 1397-1404.
[25] 李卉, 马云, 马波, 高扬, 宋辉. 不同周龄小鼠GnIH的血清含量及卵巢表达研究. 农业科学研究, 2017(4): 13-15.
LI H, MA Y, MA B, GAO Y, SONG H. Study on serum content and ovarian expression of GnIH in mice of different weeks of age.2017(4): 13-15. (in Chinese)
[26] 汪瑶, 李珣, 李敏婕, 国梦婕, 雷治海. GnIH对母猪生殖调控的研究. 中国农业科学, 2014, 47(18): 3716-3724.
WANG Y, LI X, LI M J, GUO M J, LEI Z H. Study on reproductive regulation of sows by GnIH., 2014, 47(18): 3716-3724. (in Chinese)
[27] Maddineni S R, Ocón-Grove O M, Krzysik-Walker S M, Hendricks G L, Ramachandran R. Gonadotropin- inhibitory hormone (GnIH) receptor gene is expressed in the chicken ovary: potential role of GnIH in follicular maturation., 2008, 135(2): 267-274.
[28] 张蕊, 常玲玲, 穆春宇, 付胜勇, 汤青萍, 卜柱. GnIH/GnRH对鸟类排卵的调控作用. 中国家禽, 2018(1): 42-45.
ZHANG R, CHANG L l, MU C Y, FU S Y, TANG Q P, PU Z. Regulation of GnIH/GnRH on ovulatory activity in birds.2018 (1): 42-45. (in Chinese)
[29] Franois H, Langlois I, Mulsant P, Bonnet A, Benne F, Gasser F. Gonadotropins induce accumulation of insulin-like growth factor I mRNA in pig granulosa cells.1992, 86(3): 205-211.
[30] Caloni F, Ranzenigo G, Cremonesi F, Spicer L J. Effects of a trichothecene, T-2 toxin, on proliferation and steroid production by porcine granulosa cells., 2009, 54(3): 337-344.
[31] Bentley G E, Ubuka T, Mcguire N L, Chowdhury V S, Morita Y, Yano T, Hasunuma I, Binns M, Wingfield J C, Tsutsui K. Gonadotropin-inhibitory hormone and its receptor in the avian reproductive system., 2008, 156(1): 34-43.
[32] Ubuka T, Ukena K, Sharp P J, Bentley G E, Tsutsui K. Gonadotropin-inhibitory hormone inhibits gonadal development and maintenance by decreasing gonadotropin synthesis and release in male quail., 2006, 147(3): 187-194.
[33] Wang X Y, Guo G L, Zhang X, Li M, Xiao K, Hu C H, Li X. Effect of RFRP-3, the mammalian ortholog of GnIH, on the epididymis of male rats., 2018. DOI: 10. 1016/ j. theriogenology. 2018. 05. 029.
[34] Son Y L, Ubuka T, Soga T, Yamamoto K, Bentley G E, Tsutsui K. Inhibitory action of gonadotropin-inhibitory hormone on the signaling pathways induced by kisspeptin and vasoactive intestinal polypeptide in GnRH neuronal cell line, GT1-7., 2016, 30(6): 2198.
[35] Dave A, Krishna A, Tsutsui K. Direct effects of RFRP-1, a mammalian GnIH ortholog, on ovarian activities of the cyclic mouse., 2017, 252: 193-199.
[36] Wang X F, Zhou Q M, Lu Y Y, Zhang T L, Su S B. Glycyrrhetinic acid potently suppresses breast cancer invasion and metastasis by impairing the p38 MAPK-AP1 functional axis., 2015, 19 ( 5) : 577-587.
[37] Tung C L, Jian Y J, Chen J C, Wang T J, Chen W C, Zheng H Y, Chang P Y, Liao K S, Lin Y W. Curcumin downregulates p38MAPK-dependent X-ray repair cross-complement group 1( XRCC1) expression to enhance cisplatin-induced cytotoxicity in human lung cancer cells., 2016, 389 (6): 657-666.
[38] Calder M D, Watson P H, Watson A J. Culture medium, gas atmosphere and MAPK inhibition affect regulation of RNA-binding protein targets during mouse preimplantation development., 2011, 142(5): 689-698.
[39] Inagaki K, Otsuka F, Miyoshi T, Yamashita M, Takahashi M, Goto J, Suzuki J, Makino H. p38- Mitogen- activated protein kinase stimulated steroidogenesis in granulosa cell-oocyte cocultures: role of bone morphogenetic proteins 2 and 4., 2009, 150(4): 1921-1930.
[40] Nikoletopoulou V, Markaki M, Palikaras K, Tavernarakis N. Crosstalk between apoptosis, necrosis and autophagy., 2013, 1833(12): 3448-3459.
Effects of GnIH on Autophagy and Apoptosis of Porcine Ovarian Granulosa Cells via p38MAPK Signaling Pathway
Zhang Xin, Huo KongLin, Song XingXing, Zhang DuoNi, Hu Wen, Hu ChuanHuo , LI Xun
(College of Animal Science and Technology, Guangxi University, Nanning 530004)
【】Studies have shown that autophagy and apoptosis restrict each other. As one of the main regulatory pathways of apoptosis, p38MAPK signaling pathway also has the dual effects of promoting and inhibiting autophagy. It has been proved that gonadotropin inhibitory hormone (gonadotropin-inhibitory hormone, GnIH) has effects on autophagy and apoptosis, but the mechanism of action is not clear. This experiment was conducted to study the effects of GnIH on autophagy and apoptosis of porcine ovarian granulosa cells via p38MAPK signaling pathway and its mechanism. 【】Oval granulosa cells were extracted from pig ovaries and cultured. To explore the best time of GnIH on p38MAPK signaling pathway, according to the time gradient of incubation GnIH (0, 10, 30, 60, and 90 min), Western blot was used to detect the protein expression of p38 and p-p38 in pGCs. To verify the effect of GnIH on p38MAPK signaling pathway, the cells were divided into 4 groups (control, GnIH, p38 activating agent (U-46619), and U-46619 +GnIH), Western blot was used to detect the protein expression of p38 and p-p38. To investigate the effects of different concentrations of GnIH on autophagy and apoptosis: the cells were divided into 5 groups ( control, 10-6mol·L-1GnIH, 10-8mol·L-1GnIH, 10-10mol·L-1GnIH, and 10-12mol·L-1GnIH), Western blot was used to detect the protein expression of autophagy and apoptosis. To verify the effects of different concentrations of GnIH on autophagy and apoptosis through p38 signaling pathway: the cells were divided into 6 groups (control, U-46619, U-46619+10-6mol·L-1GnIH, U-46619+10-8mol·L-1GnIH, U-46619+10-10mol·L-1GnIH, and U-46619+10-12mol·L-1GnIH), Western blot was used to detect the protein expression of autophagy and apoptosis. 【】After incubation with GnIH for 10 min, the protein expression of p38 and p-p38 was significantly decreased (<0.05). The results suggested that the optimum action time of GnIH on p38MAPK signaling pathway was 10 min; U-46619 significantly promoted the phosphorylation of p38 in pGCs (<0.05), while GnIH significantly inhibited p38 phosphorylation of pGCs (<0.05). The results suggested that U-46619 activated the p38MAPK signaling pathway, and GnIH inhibited the activation of p38MAPK signaling pathway; When the concentration of GnIH was 10-6mol·L-1, the autophagy and apoptosis of pGCs increased significantly (<0.05). With the decrease of GnIH concentration, the autophagy level of pGCs increased gradually (<0.05), while the apoptosis level of pGCs decreased gradually (<0.05). The results suggest that high concentration of GnIH promote autophagy and apoptosis. With the decrease of GnIH concentration, the autophagy level increased gradually, while the apoptosis level decreased gradually. After adding U-46619, GnIH significantly upregulated the autophagy of pGCs and down-regulated the apoptosis of pGCs (<0.05), which suggested that different concentrations of GnIH affected the autophagy and apoptosis of pGCs through p38MAPK signaling pathway. 【】GnIH might up-regulate the autophagy of pGCs and reduce the apoptosis of pGCs by inhibiting the activation of p38MAPK signaling pathway.
autophagy; apoptosis; GnIH; p38MAPK; porcine; ovarian granulosa cells

10.3864/j.issn.0578-1752.2020.09.016
2018-11-30;
2020-03-11
国家自然科学基金青年基金项目(31402151)、广西自然科学基金面上项目(2017GXNSFAA198086)、广西自然科学基金青年基金项目(2015GXNSFBA139077)
张鑫,Tel:15676119603;E-mail:2507854337@qq.com。通信作者李珣,Tel:15578947781;E-mail:lixun198@163.com。通信作者胡传活,E-mail:hch64815@gxu.edu.cn
(责任编辑 林鉴非)
猜你喜欢
中国畜牧杂志(2022年11期)2022-11-17
中国畜牧兽医(2022年8期)2022-08-23
当代畜禽养殖业(2022年3期)2022-07-13
北京航空航天大学学报(2022年6期)2022-07-02
人人健康(2021年21期)2021-12-09
集装箱化(2021年1期)2021-04-12
天津医科大学学报(2021年1期)2021-01-26
湖北农业科学(2020年22期)2020-12-18
保健与生活(2020年18期)2020-09-26
中国畜牧杂志(2020年3期)2020-03-17
