
大腹园蛛管状腺丝重组蛋白的克隆表达及纺丝
2020-04-27 13:19:58汪双祝王康康田露阳
工业微生物 2020年2期
汪双祝,温 睿,王康康,田露阳,孟 清,林 瑛
东华大学化学化工与生物工程学院, 上海 201620
蜘蛛丝是一种天然的高分子蛋白纤维,并因其优异的性能及巨大的应用前景而受到科研人员的广泛关注,蜘蛛丝在生物医学上有十分优良的应用前景,包括可以作为组织工程支架,药物运输载体和神经导管等。一只成熟的雌性圆网蜘蛛具有7种不同的丝腺体,并且能够纺出6种不同的丝纤维以及一种粘液蛋白[1]。这些不同类型的丝纤维各自具有独特的机械性能并具有不同的用途[1]。目前已有多条蜘蛛丝的全基因编码序列的报道,还有研究对蛛丝蛋白的部分结构域进行了高级结构解析[2-4]。如何简便、低成本地合成仿生蜘蛛丝纤维一直是国内外学者研究的热点,而蛛丝蛋白成丝机理的研究是蛛丝蛋白仿生应用的关键问题。20世纪初针对蛛丝成丝机理提出了两种假说,分别是“液晶态模型(liquid crytal theory)”[5]和“小球模型(micelle theory)[6]”,但都不能完整的阐述其组装成丝的过程。
七种蛛丝纤维中拖丝因其高强度受到研究者青睐,关于管状腺丝的研究相对较少。管状腺丝主要由TuSp1蛋白构成,是蜘蛛卵袋外层丝的主要成分,用于保护蜘蛛卵免受外界环境侵害,其强度可达1 GPa,且硬度在几种蛛丝纤维中是最强的[6-8],具有很好的仿生应用前景。关于大腹园蛛TuSp1的全基因序列在2017才被报道出来[9]。与其他蛛丝蛋白类似,大腹园蛛TuSp1蛋白可分为三个部分:非重复氨基末端结构域(NT),非重复羧基末端结构域(CT)以及位于蛋白序列中央的重复区(Rp)[10-14];并且相比于其他蛛丝蛋白,TuSp1的基因序列较小,其包含9个重复单元,序列中富含丙氨酸与丝氨酸[9]。但目前仍缺乏对大腹园蛛TuSp1结构与功能的研究,关于大腹园蛛TuSp1的蛋白表达及是否能自组装成丝的问题也没有相关报道。
为了使管状腺蛛丝蛋白TuSp1得到更好的仿生应用,本文采用PCR技术从已获取的大腹园蛛TuSp1的完整基因序列中扩增出其NT,CT以及Rp三种结构域序列,成功构建出大腹园蛛重组TuSp1 NT-Rp-CT克隆,并对其进行蛋白表达,纯化。通过对不同pH条件下重组蛋白液圆二色谱测试来初步分析其二级结构在不同pH下的变化。并进一步对重组蛋白冻干、重溶及湿法纺丝,进行红外光谱测试来探讨重组蛋白丝纤维成形后二级结构的变化趋势。最后用扫描电镜观察了大腹园蛛天然包卵丝纤维(TuSp1)、重组蛋白冻干形成的絮状物及湿纺后的丝纤维的具体形态特征,以探索其自组装成丝的能力。这些研究都为TuSp1成丝机理以及人工蛛丝纤维制备提供了有力的数据支持与理论依据。
1 材料与方法
1.1 材料
质粒扩增宿主细胞大肠杆菌T1与表达宿主细胞BL21(DE3)购于北京全式金生物技术有限公司;质粒提取、PCR产物回收等用于克隆表达的试剂盒购于生工生物工程(上海)股份有限公司;TaqDNA酶聚合酶购于南京诺唯赞生物科技有限公司;DNA限制性内切酶与DNA连接酶购于Thermo Fisher Scientific公司(美国);测序在上海睿迪生物有限公司进行。蛋白纯化相关试剂购于Qiagen公司(德国);Western blot相关试剂购于Invitrogen 公司(美国)。
1.2 重组蛛丝蛋白克隆的构建与表达
1.2.1克隆构建
根据获取的大腹园蛛A.v. TuSp1的完整基因序列,设计引物分别克隆A.v. TuSp1的NT-R、CT。在重复区编码序列反向PCR引物和CT端正向引物引入NdeI酶切位点,NT端正向引物引入NheI酶切位点,CT端反向引物引入Xho I酶切位点,引物序列如表1。以提取的大腹园蛛基因组DNA为模板,NT端正向引物PNTF与R反向引物PR1进行PCR扩增,获得片段NT-R、CT;以CT端正、反向引物PCTF和PCTR可扩增出CT片段。PCR反应体系均为50 μL,具体为:基因组DNA模板1 μL,正反向引物各2 μL,混合物2×TaqPlus Master Mix 25 μL,最后用ddH2O补齐至50 μL。PCR扩增程序为:95 ℃预变性5 min,95 ℃变性30 s,62 ℃退火30 s,72 ℃延伸1 min,循环30次,72 ℃ 补平10 min,4 ℃保存。之后,对PCR产物进行琼脂糖凝胶电泳检测并使用试剂盒进行胶回收。
对PCR反应回收得到的片段产物NT-R、CT以及表达载体质粒ppSUMO进行酶切,室温下连接1 h,获取完整的重组表达质粒TuSp1 NT-Rp-CT。

表1 Mini-TuSp1 PCR引物
注: 下划线标记碱基序列依次为NheI、NdeI、NdeI和XhoI酶切识别序列。
1.2.2TuSp1 NT-Rp-CT重组蛋白的表达与纯化
重组质粒经转化导入到大肠杆菌表达菌株 BL21( DE3)感受态细胞,将其涂布于平板上进行过夜培养。挑平板上单克隆接种于10 mL LB培养基中(Amp抗性,浓度为100 μg /mL)。随后过夜培养物按1∶100比例转接扩大培养至1 L LB 培养基中,37 ℃、200 r/min培养至OD600为0.6~0.8时,加入终浓度0. 3 mmol /L IPTG,16 ℃、180 r/min,过夜诱导。
5 000 r /min、4 ℃、15 min离心收集过夜诱导表达的菌体,加入50 mL 20 mmol /L Tris-HCl(pH 8.0)进行重悬并置于冰上,随后利用JN-3000 plus型高压细胞破碎机(100 MPa)进行菌体破碎,重复破碎3次后将其用50 mL离心管收集。分别取样上清与沉淀,进行SDS-PAGE检测,最终发现重组蛋白主要表达在沉淀里。然后弃上清,收集沉淀,在其中加入50 mL 含6 mol/L的盐酸胍裂解缓冲液进行重悬,待沉淀大致溶解后,采用350 W超声破碎的方式进一步混匀(超声5 s,间隔7 s,重复99次)。重悬混匀后,10 000 r/min、4 ℃、30 min离心收集上清进行蛋白纯化。因重组蛋白带有6×His标签,所以实验中使用镍柱亲和纯化目的蛋白,具体纯化步骤参考Qiagen镍柱使用手册。
1.2.3重组蛋白的湿法纺丝
将纯化透析后的重组蛋白放于冻干机中冷冻过夜,冻干后的重组蛋白按20%比例溶于六氟异丙醇中,溶解完全后对其进行高速离心以除去纺丝液内的不溶杂质。纺丝凝固浴选取100%甲醇,纺丝槽的温度设定为25 ℃。使用微量匀速泵将纺丝液以恒定流速推入凝固浴中,并立即用镊子夹住已形成的丝纤维开始拉伸,经过一段距离后拉出凝固浴,缠绕于马达上的线轴上。不断调整线轴的角速度直至丝纤维的直径变得均匀且丝纤维表面呈现出较光滑的状态,收集丝纤维置于通风橱内24 h促使甲醇挥发干净。
1.3 二级结构测试
1.3.1圆二色谱测试
将经镍柱纯化的目的重组蛋白透析于5 L的20 mmol/L 磷酸盐缓冲液中,再转移至浓缩管进行浓缩。测试前,先使用不同pH(7.0、6.5、6.0、5.5)的磷酸盐缓冲液进行稀释,并使蛋白溶液的终浓度保持在0.1~0.3 mg/mL,稀释后的蛋白液可用于圆二色谱测试。设置测试条件参数:设定光谱范围为190 nm~260 nm,波长的带宽(bandwith)为1 nm,波长步进(wavelength step)1 nm,平均时间(averaging time)为0.5 s,多次扫描等待时间(multi-scan wait)0 s;测量得到的CD图谱使用CD Spectroscopy Deconvolution Software(CDNN 2.1)进行平均、背景消除及平滑处理。
1.3.2红外光谱测试
对湿法纺丝后的蛋白丝纤维利用傅里叶变换红外-拉曼光谱仪(NEXUS-670,USA)进行红外光谱(ATR-FTIR)测试 ,操作严格确保室内温度且湿度控制在65%以下。最后所得的图谱用Peakfit V4.12软件处理,对位于酰胺Ⅰ带(1 700 cm-1~1 600 cm-1)的拉曼光谱二阶导算法处理后进行Gauss峰行拟合,估算子峰的位置以及个数,拟合后使残差满足r2≥ 0.999,使用Origin 8对拟合后的图谱绘图,分析子峰与蛋白质二级结构的对应关系,根据峰面积计算蛋白质二级结构的百分比例。
1.4 扫描电镜观察
对重组蛋白冻干形成的絮状物和湿纺后的丝纤维进行电镜扫描。制备样品,喷金20 s。用日立TM-1000 台式扫描电镜,采用12 kV电压,BSD模式观察并拍照。
2 结果
2.1 重组克隆的鉴定
将构建好的重组表达质粒转化到大肠杆菌T1感受态细胞,进行菌液PCR鉴定,后提取质粒送于生工生物工程有限公司进行测序。通过SnapGene软件,对测得的序列进行比对分析,发现无突变与移码现象,重组表达载体质粒构建成功。完整的重组蛛丝蛋白TuSp1 NT-Rp-CT氨基酸序列如图1所示,其中红色区域为TuSp1的NT端氨基酸序列,灰色区域为为TuSp1的重复区Rp端的氨基酸序列,黄色区域为TuSp1的CT端氨基酸序列。由于目前对重组蛛丝蛋白的纯化大多采用的是镍柱亲和纯化且6×His标签对其二级结构表征无影响[15,16],所以本次实验通过克隆构建,在重组表达质粒的NT端成功引入了6×His标签,方便蛋白表达鉴定与纯化以制备丝纤维。


图1 重组蛋白TuSp1 NT-Rp-CT的氨基酸序列
2.2 重组蛋白的表达与纯化
将构建好的重组表达质粒导入到大肠杆菌进行外源诱导表达,SDS-PAGE结果如图2(A)所示,与诱导前泳道N相比,诱导后泳道I中的外源重组蛋白得到大量表达,并通过Western Blot进一步验证了其表达的蛋白为所要的重组蛛丝蛋白TuSp1 NT-Rp-CT,大小约为50 600,且与上清泳道S相比,大部分蛋白表达在沉淀里。所以在后续蛋白纯化实验中通过加入50 mL 6 mol/L的盐酸胍使蛋白质充分变性,促进重组蛋白的溶解。
经盐酸胍溶解后的重组蛋白大部分溶于上清中,采用镍柱进行亲和纯化,蛋白纯化后的结果如图2(B)所示,经Image J软件处理分析,纯化后的重组蛋白纯度高达90%。收集纯化后的蛋白液,透析去除尿素,4 ℃储存,以用于二级结构分析。
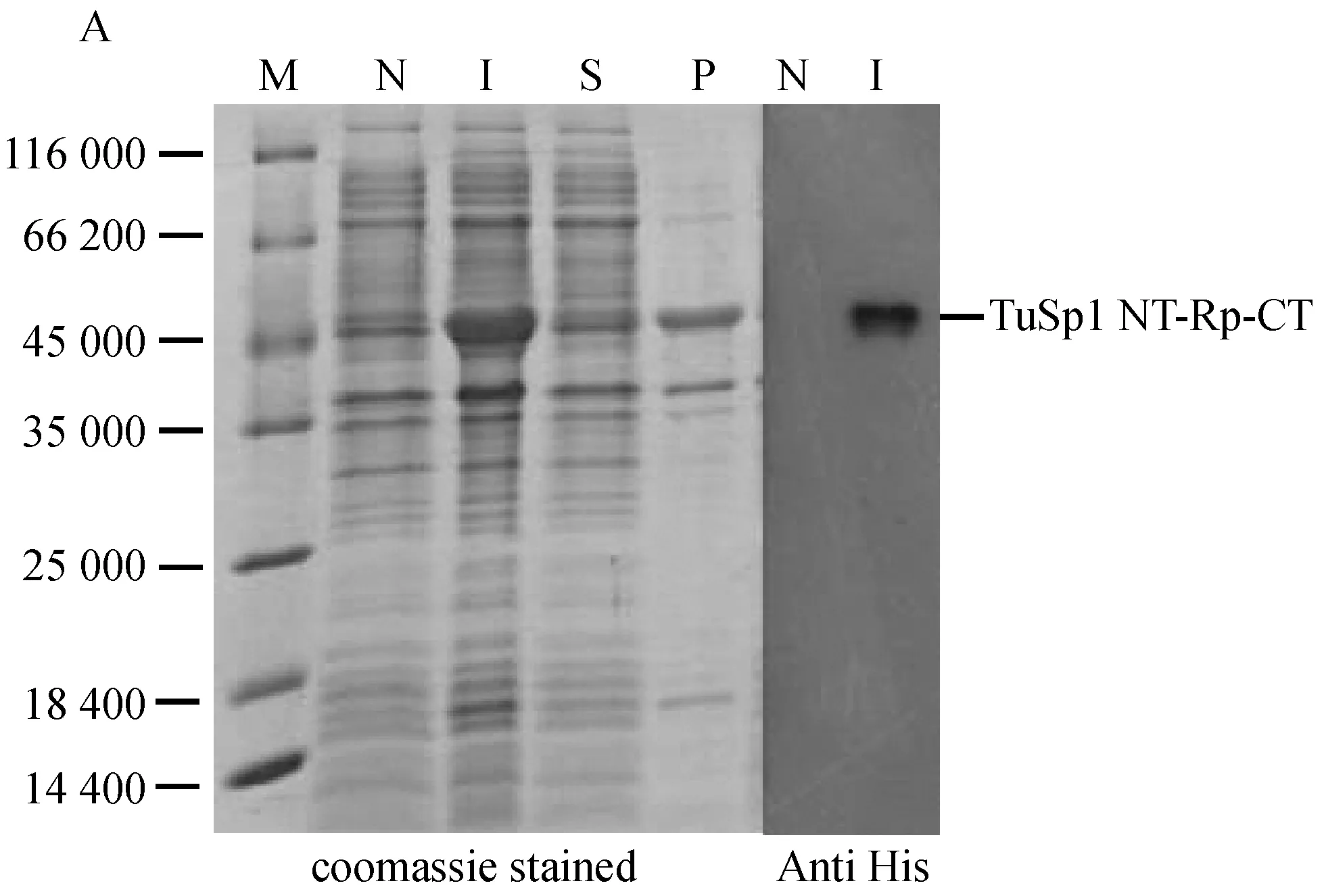

M: 蛋白质marker;N: 诱导前;I: 诱导后;S:上清; P: 沉淀;E: 纯化后的重组蛋白
图2 重组蛋白TuSp1 NT-Rp-CT的表达与纯化
2.3 重组蛛丝蛋白的二级结构
CD图谱表明,不同pH条件下,重组蛋白均呈现出α-螺旋的特征吸收峰,即在192 nm处为正峰,在208 nm和222 nm处为负峰,但不同pH条件下CD曲线几乎重叠,如图3所示,说明TuSp1 NT-Rp-CT重组蛋白均以α-螺旋构象的形式稳定存在。
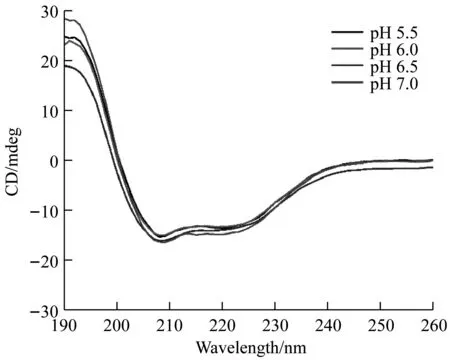
图3 重组蛛丝蛋白不同pH的CD图谱
对湿纺成丝后的蛋白纤维进行红外光谱测试,如图4所示,在(1 600~1 700) cm-1之间有吸收峰,位于酰胺Ⅰ带区域,其中峰位在(1 650~1 658) cm-1的为α-螺旋,(1 605~1 639) cm-1与(1 690~1 695) cm-1的为β-折叠,(1 659~1 689) cm-1的为β-转角,(1 640~1 649) cm-1的为无规卷曲。采用基线校正,Gaussian去卷积,二阶导数拟合对红外光谱进行处理,根据峰面积计算各二级结构的比例,结果显示(表2),β-折叠所占含量最高,且与重组蛋白液CD图谱相比,其α-螺旋有所下降,已不是主要二级结构组分。
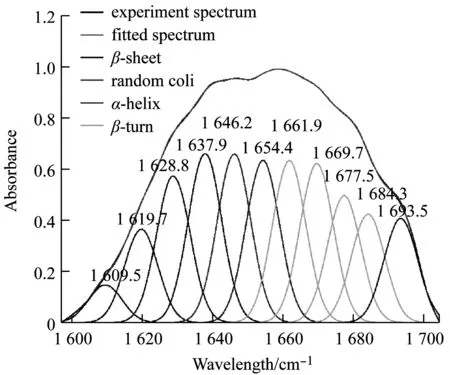
图4 重组蛛丝蛋白的红外光谱图谱

表2 重组蛋白丝纤维的二级结构统计
2.4 重组蛛丝蛋白的扫描电镜表面观察
采用扫描电镜来观察重组蛛丝蛋白TuSp1 NT-Rp-CT的丝纤维形态,电镜结果如图5所示。其中图A与图B分别展示的是电镜倍数为1 000×与5 000×下的天然包卵丝(TuSp1)纤维;图C与图D展示的是电镜倍数为1 000×与5 000×下湿纺后的重组蛛丝蛋白纤维。经对比发现,湿纺形成的重组蛛丝蛋白纤维表面整体光滑,略显粗糙且具有细小孔径,直径约24.6 μm,比天然蛛丝粗但整体较均匀。

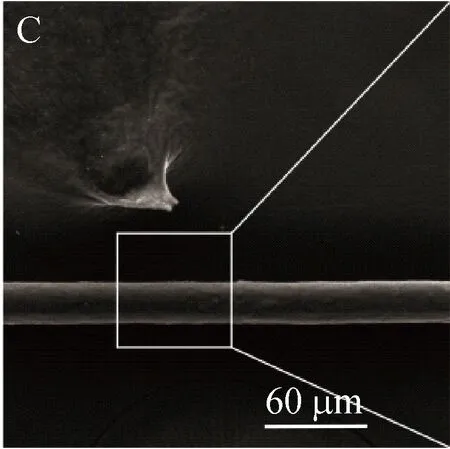

图E与图F展示的是电镜倍数为1 000×与5 000×下冻干后重组蛋白自组装丝纤维,其结果说明重组蛛丝蛋白在进行冻干过程中不施加外力的情况下具有自组装成丝的能力,且丝纤维表面光滑,直径较小,形态更接近于天然蛛丝纤维。
3 结果与讨论
蛛丝蛋白的NT结构域、CT结构域和重复区在成丝过程中起到不同的作用,有文献报道蛛丝蛋白中影响丝纤维机械性能的主要为重复区的数量和模块不同,而NT与CT作为大多数蛛丝蛋白的保守结构域对成丝也具有不同的调节作用,所以对蛛丝蛋白三种基本结构的探索是阐述其成丝机制的必要条件[17-21]。目前已有大腹园蛛TuSp1蛛丝蛋白的全长基因序列,并且其氨基酸序列二级结构预测显示NT结构域存在5个α-螺旋和两个半胱氨酸残基,CT结构域存在6个α-螺旋,但对大腹园蛛TuSp1蛋白还缺乏深入研究,这限制了TuSp1蛋白成丝机理的探索和仿生应用。
为解决这一问题,本研究模拟天然蛛丝结构,通过克隆构建成功获取重组蛛丝蛋白TuSp1 NT-Rp-CT。经蛋白诱导表达、SDS-PAGE检测,发现重组蛋白TuSp1 NT-Rp-CT主要表达在沉淀里,分析原因可能是重组蛋白在大肠杆菌中易于表达,且表达量很高,合成速度太快以至于没有足够的时间进行折叠,从而以不可溶的状态即包涵体的形式存在[22]。为了获取高纯度的重组蛋白进行后续实验,我们采用强变性剂盐酸胍使蛋白充分变性并溶于上清,进行镍柱亲和纯化和梯度透析复性后,最终得到了高纯度的重组蛋白液。CD图谱表明重组蛋白主要以α-螺旋构象存在,并且发现不同pH条件下的CD曲线无明显变化,说明一定范围下的pH波动对重组蛋白TuSp1 NT-Rp-CT的结构无影响,其结构改变可能发生在高级结构,需要进一步进行实验证实。但对比成丝后的蛋白丝纤维的红外光谱结果,发现其纺成丝纤维后二级结构由α-螺旋构象主要转变为β-折叠与β-转角构象。推断这种结构的变化趋势可能是由于纯化后的蛋白进行了高浓度的重溶,随后在湿纺中施加了一定的剪切力,使得其快速成为固体丝纤维,当然这其中还包含许多复杂的环境因素,有待进一步考究[5, 6]。同时在实验中获得了其湿纺前的蛋白丝纤维,说明重组蛋白具有自组装成丝的能力,并进一步对纺丝前后的蛋白丝纤维进行了电镜扫描观察,发现二者形态均接近天然蛛丝,但湿纺后的丝纤维较粗且具有细小孔径,推测可能是在湿纺过程中手法的限制及其他环境因素的影响。
综上所述,本文通过外源表达纯化成功获得了TuSp1 NT-Rp-CT重组蛋白,表达量可达70 mg/L;探究了重组蛋白在成丝过程中二级结构由α-螺旋向β-折叠转化的趋势。上述研究结果为探究大腹园蛛TuSp1蛋白结构与功能的关系及其高级结构解析奠定基础,同时对TuSp1蛋白成丝机理进行了初步探索,为其仿生应用奠定了理论基础。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04 11:40:51
生物学通报(2022年1期)2022-11-22 08:12:18
疯狂英语·新悦读(2021年4期)2021-11-24 09:35:53
南京林业大学学报(自然科学版)(2021年5期)2021-10-13 02:06:16
疯狂英语·新阅版(2021年4期)2021-08-16 03:26:02
滇池(2019年8期)2019-08-30 09:03:50
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
广西林业科学(2016年3期)2016-03-16 05:43:25
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
