
nanos1基因在日本血吸虫不同发育时期的定位
2020-04-01 04:54华梦晴邵延靖沈际佳
安徽医科大学学报 2020年2期
华梦晴,邵延靖,刘 淼,沈际佳
血吸虫病是一种危害严重的地方性疾病,我国流行的主要是日本血吸虫病。迄今为止,尚无针对日本血吸虫病的疫苗问世。吡喹酮因其疗效好、副作用少、疗程短等优点早已成为治疗和控制此病的首选药物[1]。然而在流行地区长期、反复、大规模的使用吡喹酮有可能会导致甚至加速耐药虫株的产生[2]。因此,寻找和研究生殖相关的基因和蛋白,为研制能够预防和治疗血吸虫病的新型干预手段具有重要意义。
nanos基因是母源效应基因,其编码产物是一种在羧基端具有两个保守锌指结构域的RNA结合蛋白,通常与pumilio共同发挥作用。nanos蛋白的功能最早发现于模式生物果蝇体内,它是果蝇建立前后体轴所必需的[3]。目前,在各种各样的物种体内都已发现了nanos同源物,研究[4-5]结果证实nanos蛋白是大多数生物的生殖细胞发育过程中的关键物质。这些研究进展是研究日本血吸虫nanos蛋白的重要参考,也是寻找能够干预血吸虫生殖发育靶标的重要思路。该研究通过多重序列比对和构建进化树对日本血吸虫nanos1蛋白进行系统进化分析,并通过原位杂交的方法探索nanos1在日本血吸虫不同发育时期的表达情况,以期为日本血吸虫生殖发育相关通路提供一定的研究基础。
1 材料与方法
1.1 主要材料日本血吸虫感染阳性钉螺购自湖南省血吸虫病防治研究所。新西兰兔和6~8周昆明鼠由安徽医科大学实验动物中心提供。PMD18-T、Taq DNA聚合酶、连接酶、DL 2000 DNA Maker购自大连宝生物有限公司(Takara)。DM10000、DM5000均为北京康为世纪生物科技有限公司产品。链霉素、青霉素购自意大利BBI公司。中量提取质粒试剂盒购自美国Omega 公司。DIG RNA Labeling Kit(SP6/T7)(C.I. 11 175 025 910)、pSPT19、DIG Nucleic Acid Detection kit(Cat.No.11175041910)均购自瑞士罗氏公司。其余试剂为国产分析纯。
1.2 方法
1.2.1多序列比对及系统进化分析 通过在线软件BlastP在NCBI中对日本血吸虫nanos1蛋白(CAX 69806.1)进行同源序列搜索,选取nanos同源蛋白共15条序列,利用DNAMAN 6.0和ClustalX 2.0.9软件对日本血吸虫nanos1氨基酸序列和其他物种nanos氨基酸序列进行多重序列比对。利用MAGA 5.05软件,以ML(Maximum Likelihood)法构建系统进化树,bootstrap检验值取1000。
1.2.2原位杂交
1.2.2.1探针的制备 根据日本血吸虫nanos1基因(FN314073.1)扩增及测序结果,利用在线软件BLAST选取了nanos1基因第416碱基至第657碱基、长度为242 bp的特异性序列。针对此段序列设计并合成引物,在上下游引物5′端分别加上限制性内切酶EcoR I和Hind III的酶切位点(下划线部分)。上游引物:5′-GAATTCTTTGTCTCCTGGAAATGCCTGC-′3;下游引物:5′-AAGCTTGCCTGGGCAGTATTTGAT-′3。PCR扩增目的片段,然后与PMD18-T载体连接构建nanos1-PMD18-T质粒,连接产物转化感受态XL1-BLUE大肠杆菌,在含有氨苄青霉素(ampicillin,AMP)抗性的LB固体培养基上培养14~16 h后随机挑取克隆进行鉴定。经DNA测序正确的nanos1-PMD18-T质粒和pSPT19载体同时经EcoR I和HindⅢ双酶切,并将nanos1-PMD18-T质粒的酶切产物与pSPT19骨架载体连接构建nanos1-pSPT19质粒。DNA测序正确后,Hind III或EcoR I单酶切nanos1-pSPT19质粒使其线性化,即为体外转录正义链探针或反义链探针所需的模板。利用DIG RNA Labeling Kit (SP6/T7)试剂盒进行体外转录,从而获得DIG标记的正义链或反义链RNA探针。探针制备成功后分装、存放于-80 ℃备用。
1.2.2.2日本血吸虫的收集 按常规方法将日本血吸虫阳性钉螺放在含去氯水的烧杯中逸尾蚴3 h,用腹部贴片法感染新西兰兔,感染后18、36和42 d从其肠系膜静脉获取日本血吸虫。为保证虫体的完整,用细毛笔在冰上分离雌雄虫并置于RNA Later中-80 ℃保存。
1.2.2.3日本血吸虫整体原位杂交 将分离的雌雄虫分别放入底部带网(75 μm目)的EP管内,4%多聚甲醛(paraformaldehyde,PFA)常温条件下固定30 min。将雄虫用0.1% SDS溶液(溶于50 mmol/L DTT、1% NP-40)在37 ℃条件下处理10 min,雌虫不做此步处理。梯度甲醇脱水后置于6% H2O2溶液(用甲醇稀释)中在日光灯下漂白20 h,以去除虫体肠腔内的底色。梯度甲醇水化后置于0.1% SDS溶液(溶于PBSTx)内孵育10 min。蛋白酶K(1 μg/ml,溶于PBST)作透膜处理后用甘氨酸溶液(2 mg/ml,溶于PBSTx)终止蛋白酶K反应,冰上放置5 min。用PBSTx洗2次,4%PFA后固定10 min。将雌雄虫移入12孔板内,加入预杂交液56 ℃预杂交2 h;然后加入含有正义或反义探针的杂交液,56 ℃杂交20 h,探针的终浓度为5 ng/ml。杂交后用柠檬酸钠缓冲液(saline sodium citrate,SSC)梯度洗脱未结合的探针,然后室温下马来酸缓冲液(含0.1 mol/L 马来酸、0.15 mol/L NaCl,pH 7.5)洗2次。封闭液(用马来酸缓冲液稀释)室温下封闭2 h,anti-dig-AP抗体 4 ℃孵育过夜,抗体清洗液洗脱未结合的抗体,然后加入NBT/BCIP避光显色。显色结束后PBSTx彻底清洗,4% PFA固定10 min后再用PBSTx洗2次,无水乙醇洗20 min以去除浮色,50%乙醇和PBSTx各洗1次,最后将虫体保存于80%甘油中,正置显微镜(Olympus DP73,日本)下观察并采集图像。
2 结果
2.1 生物信息学分析日本血吸虫nanos1蛋白的氨基酸序列与其他物种nanos同源物的一致性分别为曼氏血吸虫nanos1(XP_018652467.1)23%、 曼氏血吸虫nanos2(CCD79086.1)46%、人nanos1(NP_955631.1)16%、人nanos2(NP_001025032.1)16%、日本三角涡虫nanos(BAD88623.1)21%、鼠nanos1(NP_848508.2)22%、鼠nanos2(NP_918953.2)15%、日本血吸虫SJCHGC03345蛋白(AAW27569.1)16%、埃及血吸虫nanos(XP_012794143.1)21%、地中海涡虫nanos(ABO52809.1)20%。见图1。系统进化树表明日本血吸虫nanos1蛋白最终与曼氏血吸虫nanos2聚为一支,亲缘关系最近。见图2。
2.2 制备探针所需片段的扩增以及重组质粒的鉴定以日本血吸虫成虫cDNA为模板,PCR扩增后目的条带位于250 bp左右,序列与测序结果一致。重组质粒nanos1-PMD18-T、nanos1-pSPT19经HindIII和EcoR1双酶切鉴定,均得到大小约为250 bp的片段,酶切图谱与预期一致,证明上述2个重组质粒构建成功。见图3。
2.3 体外转录RNA探针的鉴定单酶切nanos1-pSPT19质粒使其成为线性化模板,体外转录nanos1基因正义或反义探针。转录产物经普通的1%琼脂糖凝胶电泳鉴定,得到大小约为250 bp的片段,表明DIG-RNA探针体外转录成功。见图4。
2.4nanos1基因在日本血吸虫的定位对感染后第18、24和42天3个发育阶段的日本血吸虫雌雄虫分别设计了正义探针组和反义探针组进行原位杂交实验。正义探针和反义探针杂交后,加入显色底物5-溴-4-氯-3-吲哚酚磷酸盐(5-bromo-4-chloro-3-indolyl-phosphate,BCIP)和氮蓝四唑盐(Nitro-Blue-Tetrazolium,NBT),阳性位置显蓝紫采用DNAMAN 6.0和ClustalX 2.0.9软件进行不同物种nanos蛋白氨基酸序列多重比对。nanos蛋白结构域用黑色下划线标示,红色框内是各物种间保守的氨基酸残基CCHC,它们组成2个连续的高度保守的锌指结构域,深色阴影部分表示所选物种中均保守的氨基酸残基色。nanos1基因高表达于第24、42天雌虫的卵巢和卵黄腺,第18天雌虫高表达于卵巢,而各个发育阶段的雄虫均未观察到明显阳性信号。感染后第18天血吸虫原位杂交结果见图5,第24天见图6,第42天见图7。

图1 日本血吸虫nanos1氨基酸序列与其他物种的nanos同源物氨基酸序列多重比对
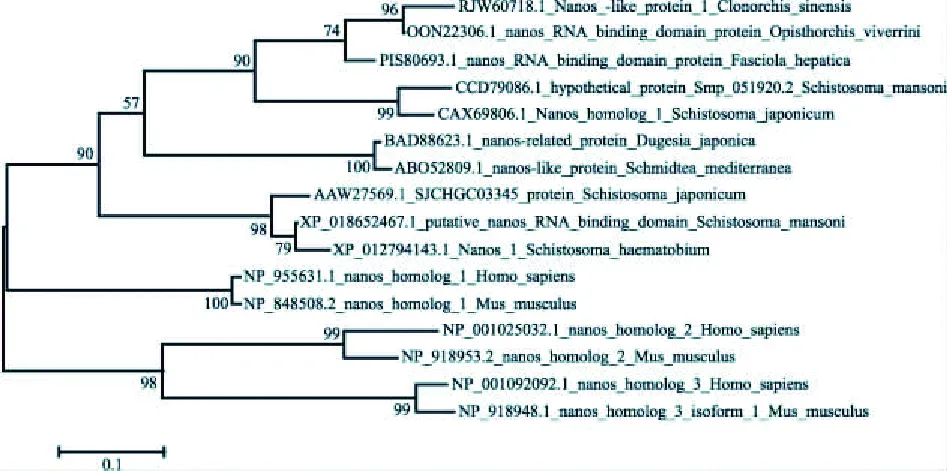
图2 基于不同物种nanos蛋白序列构建的系统进化树
采用MAGA 5.05构建进化树。日本血吸虫nanos1在进化树中的位置被“★”标记
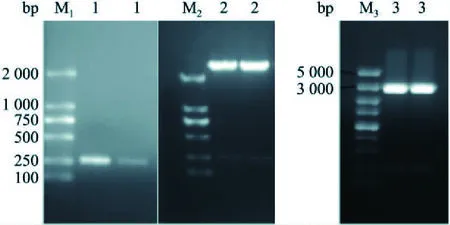
图3 PCR产物及重组质粒双酶切鉴定M1、M2: DM2000 DNA Maker;M3: DM5000 DNA Maker;1: nanos1 PCR产物;2: nanos1-PMD18-T双酶切产物;3: nanos1-pSPT19双酶切产物
3 讨论
目前研究[4-5]认为nanos基因对生殖细胞的发育和维持是必不可少的。在扁形动物真涡虫中nanosmRNA在发育、再生和成熟的睾丸和卵巢中都能检测到,RNA干扰实验[6]表明nanos缺失后会阻断真涡虫无性和有性生殖细胞的正常发育及再生。秀丽隐杆线虫有3个nanos同源基因,Nos-1和Nos-2调控原始生殖细胞的分裂增殖和存活,如果没有这两个基因,生殖细胞在第2个幼虫期结束时停止增殖,并以部分依赖于凋亡基因ced-4的方式死亡[7]。nanos-3与FBF蛋白相互作用,通过抑制fem-3 mRNA来实现雌雄同体从精子发生到卵子发生的转换[8]。在小鼠基因组中已鉴定出3个nanos同源基因,nanos-1基因敲除小鼠可正常发育,没有发现任何可检测到的异常[9]。nanos-2缺失后,虽然雌性小鼠仍然保持正常的生殖能力,但是雄性小鼠的睾丸萎缩,并表现出不育症状[10]。缺失nanos-3的雌雄小鼠均丧失生育能力,生殖腺内检测不到生殖细胞[11]。非洲爪蟾nanos1主要通过翻译抑制作用维持原始生殖细胞的发育,nanos1的缺失将导致原始生殖细胞显著减少以及生殖腺中的生殖细胞丢失[12]。这些研究进展是研究日本血吸虫nanos蛋白的重要参考,也是寻找能够干预血吸虫生殖发育靶标的重要思路。
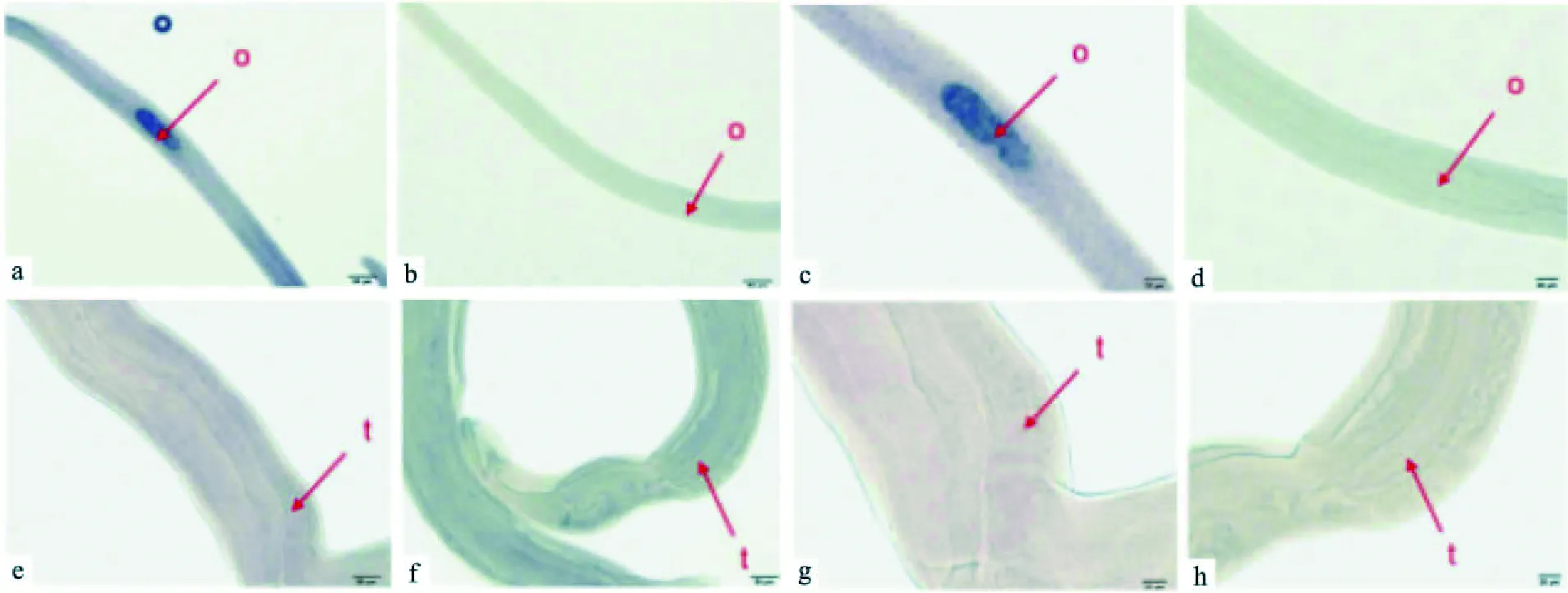
图5 感染后第18天日本血吸虫的原位杂交a、c、e、g:nanos1基因反义探针组;b、d、f、h:nanos1基因正义探针组;o:卵巢;t:睾丸

图6 感染后第24天日本血吸虫的原位杂交a、c、e、g:nanos1基因反义探针组;b、d、f、h:nanos1基因正义探针组;o:卵巢;t:睾丸;v:卵黄腺
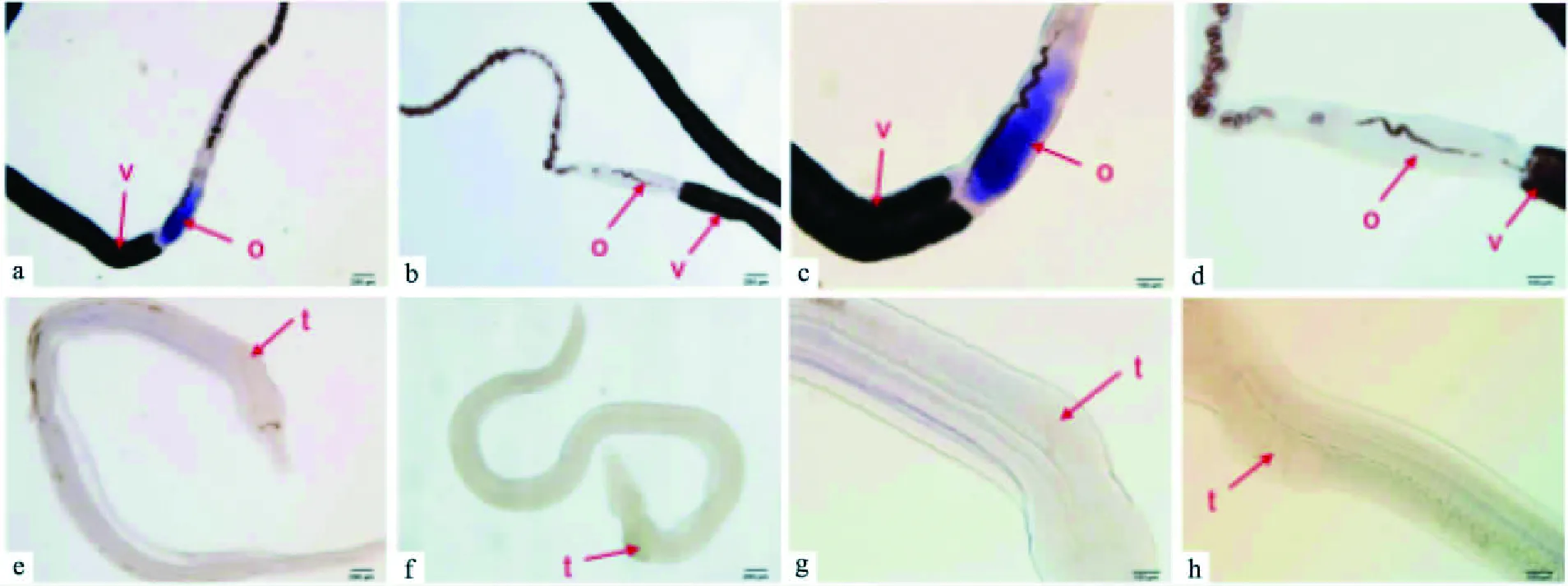
图7 感染后第42天日本血吸虫的原位杂交a、c、e、g:nanos1基因反义探针组;b、d、f、h:nanos1基因正义探针组;o:卵巢;t:睾丸;v:卵黄腺
通过对日本血吸虫基因组分析,目前已发现日本血吸虫存在两个nanos同源物,nanos1和SJCH-GC03345蛋白。本课题的研究对象是日本血吸虫nanos1,从不同物种的nanos蛋白氨基酸序列进行多重序列比对的结果来看,nanos蛋白在不同物种间同源性较低,氨基端的氨基酸序列差异很大,呈现多样性,但是在它们的羧基端都有两个高度保守且连续的C-C-H-C特异锌指结构域,符合nanos蛋白的特征。同时,可以观察到日本血吸虫nanos1蛋白与曼氏血吸虫nanos2一致性最高,达到46%。系统进化树上该蛋白最终也是与曼氏血吸虫nanos2聚为一支,亲缘关系最近。曼氏血吸虫nanos2表达于孢蚴和成虫生殖细胞内,与生殖细胞的维持和增殖相关[13],也有研究[14]报道曼氏血吸虫nanos2高表达于卵黄腺的卵黄细胞,其功能可能与性腺发育相关,由此推测日本血吸虫nanos1也是生殖相关蛋白。感染后第18、24和42天3个发育阶段的日本血吸虫雌雄虫整体原位杂交结果表明nanos1基因在雌虫的卵巢和卵黄腺高表达,而各个发育阶段的雄虫均未见明显阳性信号。但是以雄虫cDNA为模板进行普通PCR及Q-PCR均能扩增出nanos1基因片段,说明日本血吸虫雄虫同样表达nanos1基因。这就更进一步说明nanos1参与了日本血吸虫的生殖发育过程。对于雄虫原位杂交实验,课题组虽然多次调整了透膜处理时间、抗体浓度等条件,仍然没有观察到特异性杂交信号,认为其可能的原因是nanos1基因在雄虫体内的拷贝数较低或者表达丰度较低,达不到原位杂交方法检测的敏感度[15]。
综上所述,日本血吸虫nanos1基因高表达于雌虫的卵巢和卵黄腺,表明该基因参与了日本血吸虫生殖器官的发育过程,为进一步研究日本血吸虫nanos1蛋白的功能提供了线索,但是其在生殖发育相关通路中扮演的具体作用有待进一步研究。
猜你喜欢
今日健康(2022年3期)2022-11-21
保健文汇(2022年8期)2022-08-27
中国人兽共患病学报(2022年2期)2022-03-18
小学生学习指导·低年级(2021年6期)2021-09-10
考试与评价·七年级版(2021年4期)2021-08-14
分子影像学杂志(2021年2期)2021-05-25
河北渔业(2019年7期)2019-08-27
小学阅读指南·低年级版(2018年5期)2018-11-02
家庭用药(2018年3期)2018-03-23
科学大众·小诺贝尔(2018年12期)2018-02-22
