
硫化氢在植物中抵御非生物胁迫机制的研究进展
2020-03-16 03:18李子玮陈思蒙王发展张豪洋张莉许自成
中国农业科技导报 2020年4期
李子玮, 陈思蒙, 王发展, 张豪洋, 张莉, 许自成*
(1.河南农业大学烟草学院, 郑州 450002; 2.江苏中烟工业有限责任公司, 南京 210019)
硫化氢(H2S)在常温常压下是一种无色有“臭鸡蛋”味的可燃气体,可溶于水,不含水时pH 7.4,硫化氢阴离子(HS-)与H2S的比值为3∶1[1]。H2S在亲脂溶剂中的溶解度是在水中的5倍[2],所以能自由渗透脂质膜,被认为是继一氧化碳(CO)和一氧化氮(NO)后的第3种植物内源性气体信号分子。在动物和植物体中,低浓度的H2S具有多种生理功能。在动物体中,H2S影响心血管、神经元、免疫、肾、呼吸、胃肠、生殖、肝脏和内分泌系统[3],H2S参与硫代谢营养调节,提高动物体内谷胱甘肽(glutathione, GSH)含量,保护神经系统免受氧化伤害;与蛋白质活性位点的金属中心相互作用,对人体心血管和神经功能产生积极影响[4]。在植物体中参与组成含硫防御系统,包括元素硫(S0)、H2S、GSH、植物螯合剂、各种次生代谢物和富硫蛋白等[5]。植物体对H2S浓度的变化具有一定调节功能,植物细胞中过多的H2S以气体形式释放到大气中, 引发对病原体的防御响应机制,使植物细胞免受病原菌浸染[6]。本文结合国内外最新研究进展,综述了H2S在植物体中通过基因调控、改变酶活性和蛋白质的表达、硫巯基化修饰、减轻氧化应激、与信号物质的互作等抵御非生物胁迫的作用机制,并对其作用机制进行了展望,以期为气体信号分子对植物抵御非生物胁迫的调控研究提供多角度理论支持。
1 H2S产生机制
植物体既能通过叶片吸收大气中的H2S,自身也具有H2S产生的机制。Rennenberg[7]在南瓜叶片中分别添加硫酸盐、亚硫酸盐、半胱氨酸(cysteine, Cys)和SO2,也发现有H2S释放,证明使用不同的硫源存在不同的代谢途径。L-半胱氨酸脱硫酶(L-cysteine desulfhydrase, LCD)和D-半胱氨酸脱硫酶(D-cysteine desulfhydrase, DCD)是最明确的可产生内源性H2S的酶。在植物细胞的细胞核中,LCD以L-半胱氨酸(L-Cys)为底物降解产生H2S、氨(NH3)和丙酮酸。在线粒体中,DCD降解D-半胱氨酸产生H2S[8]。
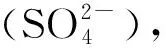
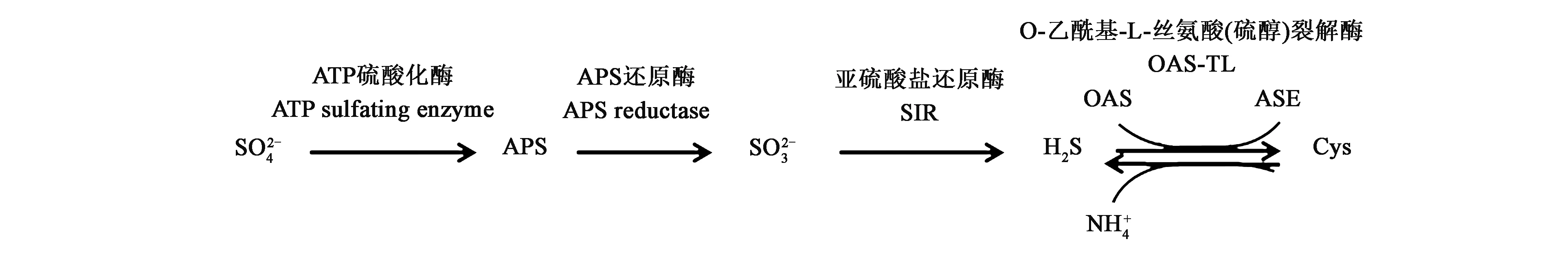
图1 植物细胞内H2S的合成[11]Fig.1 Synthesis of H2S in plant cells[11]
2 H2S抵御非生物胁迫的机制
低浓度的外源H2S可以促进植物抵御各种非生物胁迫[12]。H2S可以缓解各种非生物胁迫损害,如盐胁迫[13]、干旱和渗透胁迫[14-16]、冷胁迫[17]、热胁迫[18]和重金属胁迫[19]。
2.1 H2S通过基因调控抵御胁迫
2.1.1酶类相关基因 Fang等[20]研究表明,H2S通过调节与氨基酸和蛋白质相关的合成酶类基因,缓解Cr6+对拟南芥根系伸长的抑制作用。H2S上调半胱氨酸相关基因OASTLa、SAT1和SATS的表达水平,并延长SAT1基因和SATS基因的表达时间,促进H2S-Cys系统上调,该系统可提高植物螯合素(phytochelin,PCs)合成酶基因PCS1和PCS2的表达,增加植物螯合素的合成,减轻重金属胁迫。另一种途径同样属于基因调节,H2S上调金属硫蛋白(metallothionein,MT)相关基因MT2A的表达,减缓Cr6+胁迫。Fang等[21]的后续研究结果同样证明,H2S通过上调葡萄籽中MT3A基因和PCS基因的表达,增加与蛋白质相关的合成酶类PCs、MT等重金属螯合剂基因的表达,缓解重金属胁迫,而用H2S的合成抑制剂亚牛磺酸(hypotaurine, HT)处理此过程被阻断。Hu等[22]研究表明,丁香酚通过上调油菜根系中BrLCD基因和BrDCD基因的表达及其酶活性,内源H2S含量增加,游离Cd2+显著降低。若使用H2S的合成酶抑制剂——炔丙基甘氨酸 (propargylglycine, PAG)处理,油菜对镉的耐受性降低。
Xie等[23]研究表明, 外源H2S调节另一种合成酶类血红素氧合酶-1(HO-1) 基因的表达, 刺激内源CO产生,延缓赤霉素(gibberellin, GA) 诱导的小麦糊粉层细胞程序性死亡(programmed cell death,PCD) ,而分别加入HO-1的两种副产物一氧化碳(CO) 水溶液或胆红素时,抑制作用部分逆转,NaHS介导的PCD延缓作用消除。外源H2S诱导黄瓜HO-1转录本(CsHO1)、CsDNAJ-1和CsCDPKl/5基因的表达,CO能缓解HO-1抑制剂锌原卟啉(zinc protoporphyrin, ZnPP) 对基因表达的抑制,说明H2S在诱导黄瓜不定根发生的过程中可能是通过HO-1/CO信号系统产生的作用[24]。
H2S还可通过调控氧化还原酶类基因,缓解锌胁迫。Liu等[25]研究表明,H2S上调抗氧化酶相关基因CAT2的表达,下调氧化酶相关基因CSD、FSD和pAPX的表达,使过量的H2O2还原,减轻锌诱导的氧化应激,缓解锌胁迫。
2.1.2植物激素相关基因 H2S调节植物激素生物合成相关基因(NCED1/6、GA2ox7、IAA氧化酶、ACO、脂氧合酶)和TFs(AP2)的表达来改变激素水平[26]。茉莉酸(jasmonic acid, JA) 是高等植物的内源生长调节剂,诱导气孔关闭,作用于H2S信号的上游,H2S调节与JA生物合成相关的基因(NtLOX、NtOS、NtAOC和NtOPR) 的转录,改变激素水平提高抗旱能力[27]。H2S与乙烯相互作用,提高香蕉抗氧化基因MaAPX和抗病基因MaPAL的表达水平,提高信号转导基因MaETR、MaERS1、MaERS2和MaERS3的表达水平,说明H2S改变乙烯含量提高抗氧化等能力,通过调节自身激素水平响应非生物胁迫[28]。王韬远等[29]研究表明,在拟南芥lcd突变体中,ABA(abscisic acid)受体编码基因GCR2和GTG1的表达水平显著增强,而H2S供体NaHS处理显著降低基因表达,证明外源H2S增强了ABA诱导的气孔关闭。
2.1.3信号分子相关基因 低温胁迫诱导植物内源H2S信号的产生,这种信号可被外源施加的Ca2+和H2S增强,使Ca2+信号转导相关基因CaM、CIPK5的mRNA表达上调,增强植物对冷胁迫的抗性[30]。丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)最早发现于苜蓿,参与生长发育和各种刺激信号传导。MAPK相关基因MPK4在拟南芥的保卫细胞和烟草的远轴表皮中均有表达,参与气孔运动,H2S作用于MPK4基因下游,抑制拟南芥气孔开放,增加耐旱性[31]。杜鑫哲等[32]研究表明,H2S上调基因MPK4的表达水平,共同调控冷胁迫响应基因ICE1、CBF3、COR15A和COR15B的表达,提高拟南芥在冷胁迫下的耐受性。香蕉MAPK1基因属于MAPK基因家族,参与香蕉逆境胁迫应答[33],推测香蕉内源H2S通过上调MAPK1基因的表达抵御冷胁迫。
2.1.4RNA相关基因 miRNA (microRNA)是生物体内普遍存在的小分子RNA, 由内源基因编码于转录后,通过介导靶mRNA降解或翻译抑制来调控基因表达, 是真核细胞基因表达的重要调控机制。Shen等[34]研究表明,H2S能有效调控miRNA相关基因Mir167a、Mir167c、Mir167d、Mir393a和Mir396a的转录水平, 提高拟南芥对干旱胁迫的耐受力。
2.1.5其他与逆境相关的基因 低浓度H2S通过上调病程相关基因(MIR-393、MIR393b)的表达,改善次级代谢产物氨基酸、有机酸、糖、糖醇和芳香胺的含量,提高植物耐受力[35]。H2S上调番茄幼苗侧根生长相关的细胞循环相关基因SICDKA1、SICYCA2、CYCD3、CDKA1,及生长素响应因子基因ARF4、ARF7, 证明H2S与吲哚乙酸(indoleacetic acid, IAA)共同诱导番茄侧根的形成[36]。Garcia-mata等[37]用NaHS预处理凤仙花,气孔孔径减小,干旱相关基因(DREB2A/B、CBD29A)表达增加,表明提高了抗旱性。
2.2 H2S通过调控蛋白质表达抵御非生物胁迫
H2S信号分子调控冷胁迫响应基因CBFs的表达,基因CBF调控防冻蛋白COR (cold response) 的表达,进而增加植物对冷胁迫的抗性[37]。H2S通过调节生物钟相关基因CCA1和PRR9的表达,进而调控下游靶标CBFs基因,提高防冻蛋白活性,增加拟南芥对冷胁迫的耐受性。H2S促进与锌吸收有关的重要转运体ZRT、IRT序列的表达,抑制编码ZIP家族转运蛋白,下调对锌的吸收[38]。
2.3 H2S通过改变酶活性抵御非生物胁迫
NaHS处理可提高玉米幼苗中海藻糖-6-磷酸酸酶(treha-lose-6-phosphate phosphatase, TPP)的活性, 从而提高海藻糖的含量增强耐热性[39]。NaHS供体提高Delta(1)-吡啶-5-羧酸合成酶活性,降低脯氨酸脱氢酶活性诱导玉米幼苗脯氨酸含量的增加,减轻丙二醛(malondialdehyde, MDA)的积累,提高热胁迫下玉米幼苗的成活率[40]。外源H2S提高β-淀粉酶活性,促进黄瓜种子的萌发[41]。三磷酸腺苷酶(adenosine-triphosphate, V-ATPase)是液泡中产生质子梯度的关键酶,镉胁迫下酶的活性受到抑制,外源H2S促进ATP水解和质子运输,对V-ATPase的合成产生积极影响,恢复液泡膜内外质子梯度浓度[42]。Dai等[43]研究表明,低浓度的NaHS降低金属胁迫下小麦叶绿体中MDA和过氧化氢(H2O2)的含量,浓度为0.8 mmol·L-1时抗氧化酶活性升高的效果最明显。
H2S对包括重金属毒性在内的不同非生物胁迫诱导的氧化应激具有重要作用。H2S增加基本元素氮(N)、磷(P)和钾(K)的吸收,平衡镁(Mg)、锰(Mn)、铜(Cu)、铁(Fe)等矿物质营养元素。H2S与根瘤菌对减少苜蓿中重金属的积累具有协同作用[44],增加植物营养元素的吸收,与重金属元素产生竞争吸收,降低H2O2、ROS和MDA的含量,使SOD、POD、APX、CAT等抗氧化酶的活性增加,减轻氧化胁迫,增加地上和地下部干重,提高苜蓿对土壤中重金属的耐受性。
2.4 H2S通过平衡电解质抵御非生物胁迫
作物生长发育易受盐碱地的影响而减产,因此,维持植物体K+/Na+平衡对于适应高盐环境至关重要。H2S在离子稳态中发挥作用,有效降低植物在盐胁迫下的中毒症状。朱会朋[45]研究发现,50 mol·L-1NaHS处理胡杨(Populuspopularis) 明显缓解盐胁迫,H2S一方面通过降低质膜去极化,抑制K+外流,另一方面通过上调质膜H+泵,为Na+/H+跨膜逆向转运提供质子浓度梯度,有效维持植物体在盐胁迫下的离子稳态。Deng等[46]通过使用CaCl2(非选择性阳离子通道的抑制剂-NSCCs)、TEA+(一种低亲和力K+转运蛋白的抑制剂)或阿米洛利(一种对盐敏感的抑制剂-SOS1),研究证明NSCCs和SOS1通道是H2S降低小麦幼苗中Na+浓度的主要途径。表明H2S通过维持胞内外电解质平衡,抵御非生物胁迫。
2.5 H2S通过改变重金属元素的形态抵御重金属胁迫
Yang等[47]在柳树根部喷施NaHS溶液,研究发现,镉在叶片和根中总的吸收水平基本不变,其中不溶性镉的比例增加,可溶性镉的比例下降,证明外源H2S通过改变镉的物理形态降低植物中毒症状。铬存在几种氧化态,其中化学性质最稳定的两种形式是Cr3+和Cr6+。Cr6+胁迫抑制拟南芥根系伸长,推测信号分子H2S可能通过提高Cr3+/Cr6+比值,降低氧化态的重金属毒性。
铜(Cu)是植物生长发育必需的微量营养元素,参与植物细胞中电子传递链、蛋白质转运和催化酶等生化反应。但铜作为氧化还原活性的过渡金属,过量铜损伤根系表皮,减少根毛的数量和长度。植物虽能分泌苹果酸、柠檬酸等有机酸来改善铜的毒性,但植物自身无法调节过量的铜产生的毒性。低浓度NaHS预处理可促进Cu与体内游离氨基酸的接触,抵消Cu对小麦种子萌发的抑制作用[48]。
2.6 H2S通过蛋白质的硫巯基化修饰抵御非生物胁迫
硫巯基化修饰(S-sulfhydrylation)是信号分子H2S在植物体中发挥生物学活性的重要途径。H2S对 Cys残基进行硫巯基化修饰,Cys-SH基团转化为Cys-SSH过硫基团,直接调控蛋白质的活性[49]。H2S通过肌动蛋白的S-硫化作用,引起肌动蛋白细胞骨架变化,抑制肌动蛋白聚合。证明外源H2S调节肌动蛋白动力学,并影响根毛生长[50]。外源H2S促进拟南芥中APX硫巯基化,提高APX的活性[51]。
2.7 H2S通过维持GSH系统稳态抵御非生物胁迫
还原型谷胱甘肽/氧化型谷胱甘肽 (GSH/GSSG)是反映细胞氧化还原状态的重要参数,在豆科植物苜蓿中,GSSG比GSH高约17倍。Lai等[52]研究表明,随着NaCl处理浓度的升高,诱导苜蓿幼苗L-CD活性和内源H2S含量的增加,H2S作为内源调节剂有效调控抗氧化酶活性,提高GSH/GSSG比值,增强AsA-GSH循环代谢,重建氧化还原平衡。Mostofa等[53]研究表明,NaHS预处理可恢复盐胁迫下抗坏血酸(ascorbic acid, AsA) 含量的损失,并进一步提高GSH含量,维持AsA/DHA和GSH/GSSG比值;平衡矿物质元素含量,降低Na+的吸收和Na+/K+的比值,同时,内源H2S含量增加,保护叶绿素、类胡萝卜素和可溶性蛋白免受盐胁迫损伤。Upadhyaya等[54]研究表明,NaHS上调水稻植株中甘氨酸(glycine, Gly) 相关酶Gly I和Gly II的活性,进而维持GSH系统稳态,减缓细胞甲基乙二醛(methylglyoxal,MG)和ROS的毒性。Singh等[55]用砷胁迫处理豌豆幼苗,外源施加H2S提高内源NO水平,NO作为第二信使,增强AsA-GSH循环代谢,平衡氧化还原状态,减少ROS介导的大分子损伤,缓解过氧化对植物的损害。
3 H2S与其他信号物质的互作
3.1 H2S与Ca2+的互作
Ca2+作为通用的第二信使,是植物中重要的转导和调节因子。Ca2+结合蛋白感知Ca2+的快速增加并传递特异性信号,植物可通过此方法产生适应性反应。H2S与Ca2+具有协同作用,H2S部分贡献由Ca2+活化的抗氧化系统,减轻金属铬胁迫。Ca2+和CaM调控烟草细胞内源H2S的积累,通过调节抗氧化系统的活力诱发耐热性的形成[56]。
3.2 H2S与植物激素的互作
高等植物的生理活动处于复杂的信号网络中,不同通路间存在不同的相互作用,共同抵御非生物胁迫。H2S改变不同信号物质的平衡,调节植物生长和耐受性。水杨酸(salicylic acid, SA) 是一种酚类信号物质,作用于H2S上游,参与植物对逆境胁迫的响应,SA和NaHS预处理可增强玉米幼苗耐热性[57]。外源H2S作用于IAA上游,促进IAA和NO共同介导的不定根形成。
气孔开闭对控制植物气体交换起重要作用, 控制光合作用所需CO2的吸收和蒸腾作用水分的散失,H2S通过与植物激素相互作用,调节气孔关闭,增强植物的耐旱性。外源JA增强蚕豆叶片保卫细胞中LCD和DCD的活性,提高内源H2S水平,H2S合成抑制剂降低了JA诱导的气孔关闭,说明H2S参与JA诱导的气孔关闭。Scuffi等[58]研究表明,H2S作用于NO上游,缺乏内源H2S时,NO含量降低,导致ABA合成受阻,抑制气孔关闭。盐胁迫诱导信号分子H2O2产生,H2O2来源于NADPH氧化酶、细胞壁过氧化物酶和POA途径,H2O2提高H2S含量,ABA合成增加,促进蚕豆气孔关闭,证明H2S可能在H2O2的下游,参与气孔关闭信号转导,与激素的互作诱导蚕豆气孔运动[59-60]。
3.3 H2S、MAPK与NO信号分子的互作
大量研究已证实,H2S和NO在调节植物生长和发育中的相互作用。NO参与激活MAPK信号传导途径,参与植物防御,硝酸还原酶依赖性NO的产生与番茄中H2S诱导的硝酸盐胁迫耐受性有关[61]。丝裂原活化蛋白激酶(MAPK)级联是真核生物中通用的信号传导方式,在植物的许多生理和生物化学过程中起至关重要的作用。Xu等[62]从黄瓜中克隆了名为CsNMAPK的MAPK基因,证明了在硝酸盐胁迫下,H2S通过调节抗氧化酶活性参与MAPK/NO信号传导,对黄瓜幼苗起到保护作用,且H2S、MAPK与NO在黄瓜根系中具有交互作用。CsNMAPK的过表达增强了其硝酸盐胁迫耐受性。如图2所示,以NaHS作为H2S的供体,示意H2S、CsMAPK和NO参与减轻黄瓜幼苗根系氧化损伤的作用[62]。
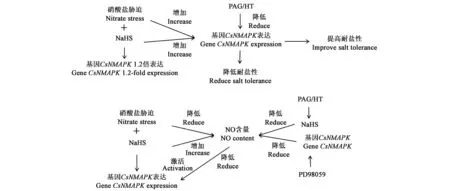
图2 H2S、MAPK与NO在黄瓜根系中抵御硝酸盐胁迫的交互作用[62]Fig.2 Interaction of H2S, MAPK and NO against nitrate stress in cucumber roots[62]
在硝酸盐胁迫下,施加H2S供体NaHS促进基因CsMAPK的表达,而用H2S抑制剂PAG和HT降低了基因CsMAPK的表达,导致黄瓜根系耐盐性降低。进一步推测,硝酸盐胁迫下NO含量较低,H2S供体NaHS提高NO含量,激活了CsMAPK的表达,而使用H2S抑制剂或基因表达抑制剂PD98059降低NO含量,下调基因CsMAPK的表达,从而降低黄瓜幼苗对硝酸盐胁迫的耐受性。
3.4 H2S与信号分子甲基乙二醛的互作
甲基乙二醛(MG)在盐胁迫中可迅速增加2~6倍,成为一种胁迫标志。Kaur等[63]通过转录组分析,用浓度为10 mmol·L-1的MG处理水稻幼苗16 h后,共1 685个基因的表达发生了1.5倍以上的变化,大部分涉及响应MG和干旱等胁迫的信号转导过程,MG响应元件(methylglyoxal-response element, MGRE) 发生10倍以上的转录变化,表明外源H2S诱导信号分子MG的表达,抵御干旱胁迫。Christou等[64]研究发现,NaHS可取代热激,通过增加草莓幼苗内源H2S的积累,同样可诱导HSP70、HSP80和HSP90基因的表达,表明外源H2S可激活MG脱毒系统,增强植物耐旱性。因此,H2S在植物信号转导过程中,与信号分子的互作触发分子机制,提高植物在逆境胁迫中的耐受能力。
在植物体中,H2S通过直接参与基因调控、改变酶活性和蛋白质的表达、硫巯基化修饰、与信号物质的互作等过程,在植物抵御非生物胁迫中发挥重要的作用,为今后更深入研究H2S的抗逆性作用机理提供理论参考,具有重要的科学意义和农业指导意义。
4 展望
H2S的功能贯穿于植物生长发育的整个过程, 尤其在植物响应非生物胁迫方面发挥至关重要的作用。目前,H2S的研究仍存在一些不足,今后的研究可以考虑从以下几个方面深入探索:①需要进一步研究H2S在植物信号转导过程中的直接靶点和上下游级联反应,还需要深入阐明H2S与其他信号分子之间互作关系网;②植物生长发育的过程取决于不同激素的比例,H2S可能是协调植物激素相互作用的中间物,其作用机理尚未明确;③结合基因技术,尝试培育抗逆作物新品种;④需要深入探索植物体内与H2S合成相关基因突变体的筛选,以及H2S荧光探针的应用。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
航空材料学报(2022年4期)2022-08-04
作物学报(2022年9期)2022-07-18
舰船科学技术(2022年11期)2022-07-15
贵州农业科学(2022年4期)2022-05-06
干旱地区农业研究(2022年1期)2022-01-28
中国土壤与肥料(2021年5期)2021-12-02
科技创新与品牌(2019年12期)2019-02-06
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
食品工业科技(2014年21期)2014-03-11
