
外源铬对延胡索生理指标及抗氧化系统变化的影响
2020-03-16 03:18盛笠程福龙余顺慧王柏慧卢增辉潘杰
中国农业科技导报 2020年4期
盛笠, 程福龙, 余顺慧, 王柏慧, 卢增辉, 潘杰*
(1.重庆三峡学院, 三峡库区水环境演变与污染防治重庆市重点实验室, 重庆 404199; 2.南京信息工程大学环境科学与工程学院, 南京 210044;3.贵州大学, 国土资源部喀斯特环境与地质灾害重点实验室, 贵阳550025;4.重庆市万州区生态环境监测站, 重庆 404100)
重金属铬(Cr)是环境污染中最危险的五类物质之一,其在土壤中大量存在时,对生态环境造成了严重的影响[1]。外源重金属Cr6+被植物体吸收后,植物体内将产生大量的活性氧(reactive oxygen, ROS),从而造成生物分子的氧化损伤[2]。因此,植物中有一套包括超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)、过氧化氢酶(catalase, CAT)、谷胱甘肽(glutathione,GSH)、抗坏血酸(ascorbate, AsA)在内的抗氧化系统来清除ROS,以保护植物细胞不受氧化损伤[3]。
已有学者研究了植物抗氧化系统在镉、铅、锌等重金属胁迫下的应激变化趋势及其作用机制[4];高扬等[5]研究发现,在Pb、Cd复合胁迫下龙葵内SOD、POD活性随重金属处理浓度的升高逐渐升高,而GSH含量逐渐减少,而其叶生物量显著减少,根茎生物量则在低浓度处理下减少不明显;何洁等[6]研究表明,随Zn胁迫含量升高,翅碱蓬发芽率、苗高、苗重均降低,SOD、POD活性也呈下降趋势,CAT活性呈先升后降趋势。延胡索(Corydalisyanhusuo)块茎是常用中药材,分布于安徽、江苏、浙江等地,目前在重庆万州地区大量引种栽培。但目前关于铬处理下延胡索抗氧化系统变化与延胡索生长、生理之间关联的研究相对较少。因此,本试验选用延胡索为受试材料,研究了铬处理下延胡索植株生理指标变化,进而探究延胡索抗氧化系统的应激变化和作用机制,以期为重金属铬对中药材毒理学研究及延胡索种植环境要求提供参考依据。
1 材料与方法
1.1 试验材料
延胡索块茎,2017年10月购于浙江省东阳市农业局,在4 ℃条件下冷藏储存备用。试验前挑选出大小基本一致的延胡索块茎120块,进行消毒处理后用于处理试验。
试验场地位于重庆市万州区重庆三峡学院种植基地(108°45′06″E,30°75′72″N)。土壤取自重庆三峡学院种植基地,将土壤样品中杂物剔除,过2 mm筛,备用。供试土壤基本性质如下:土壤类型为红土,土壤pH 6.3,有机质含量18%,全氮含量0.22%,水解氮含量12.58 mg·kg-1,速效钾含量50.34 mg·kg-1,速效磷含量11.3 mg·kg-1,Cr6+含量67.32 mg·kg-1。
1.2 试验设计
本试验依据《GB15618-2018土壤环境质量 农用地土壤污染风险管控标准(试行)》[7],采用土培试验,采用K2Cr2O7作为不同浓度外源Cr6+的处理因子,以溶液形式加入种植土壤中,设计7个K2Cr2O7处理浓度(表1),以土壤中不添加外源Cr6+处理作为对照组(CK)。每个种植盆装入8 kg供试土壤,每盆均匀播种5块延胡索块茎。按照林日长等[8]提出的技术规范进行种植管理,定期查看生长情况,实验设置3个平行。150 d后采集供试植物样本,分别测定延胡索地上部分Cr6+含量、地下部分Cr6+含量、株高、生物量、膜质过氧化物丙二醛(malonaldehyde,MDA)含量以及延胡索体内抗氧化系统指标SOD活性、POD活性、CAT活性、GSH含量和AsA含量。
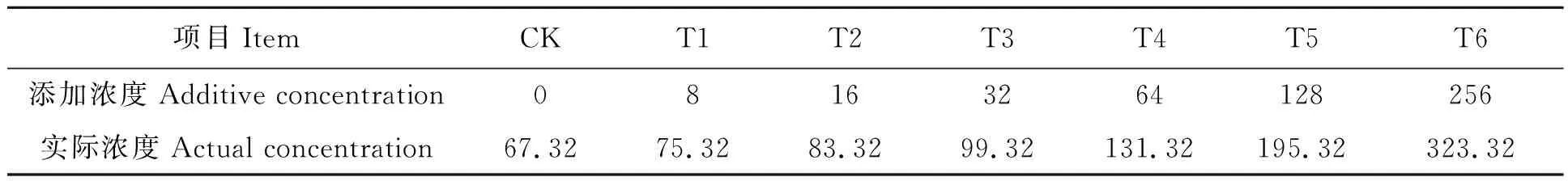
表1 各处理中土壤Cr6+浓度Table 1 Cr6+ concentration of soil under different treatments (mg·kg-1)
1.3 测定项目及方法
用刻度尺测量株高;用分析天平测量延胡索生物量;铬含量的测定采用微波消解-原子吸收分光光度法[9]进行;MDA含量的测定采用硫代巴比妥酸法进行[10];采用NBT光还原法[11]进行SOD活性的测定;CAT活性采用紫外分光光度计进行测定[11];采用愈创木酚法[11]对POD活性进行测定;GSH和AsA含量的测定方法参照Rai等[12]的方法。
1.4 数据分析与处理
用Microsoft Excel 2019处理数据、绘制图表;采用SPSS 20进行单因素方差分析。
2 结果与分析
2.1 铬处理下延胡索植株铬累积情况
图1为不同浓度Cr6+处理下,延胡索植株体内、地上和地下部分的铬含量变化情况。随着外源添加Cr6+浓度的增加,延胡索植株内、地上部分、地下部分Cr6+含量均升高;且在相同浓度Cr6+处理条件下,地上部分Cr6+含量高于地下部分Cr6+含量。同时,当对延胡素植株的处理浓度达到最高时(T6处理),延胡索体内Cr6+含量较CK增加了2倍左右,达到2 750.3 mg·kg-1DW。当土壤中Cr6+浓度在T5~T6区间时,延胡索地上部分、地下部分Cr6+含量相对于CK增加百分比最大;在T6时,地上部分、地下部分分别达到218.72%和196.24%。这说明在Cr6+处理浓度为CK~T6范围内,外源添加Cr6+浓度越高,富集含量也越高,相对于CK 的增加量也随之增加,且表现出明显的正相关关系。
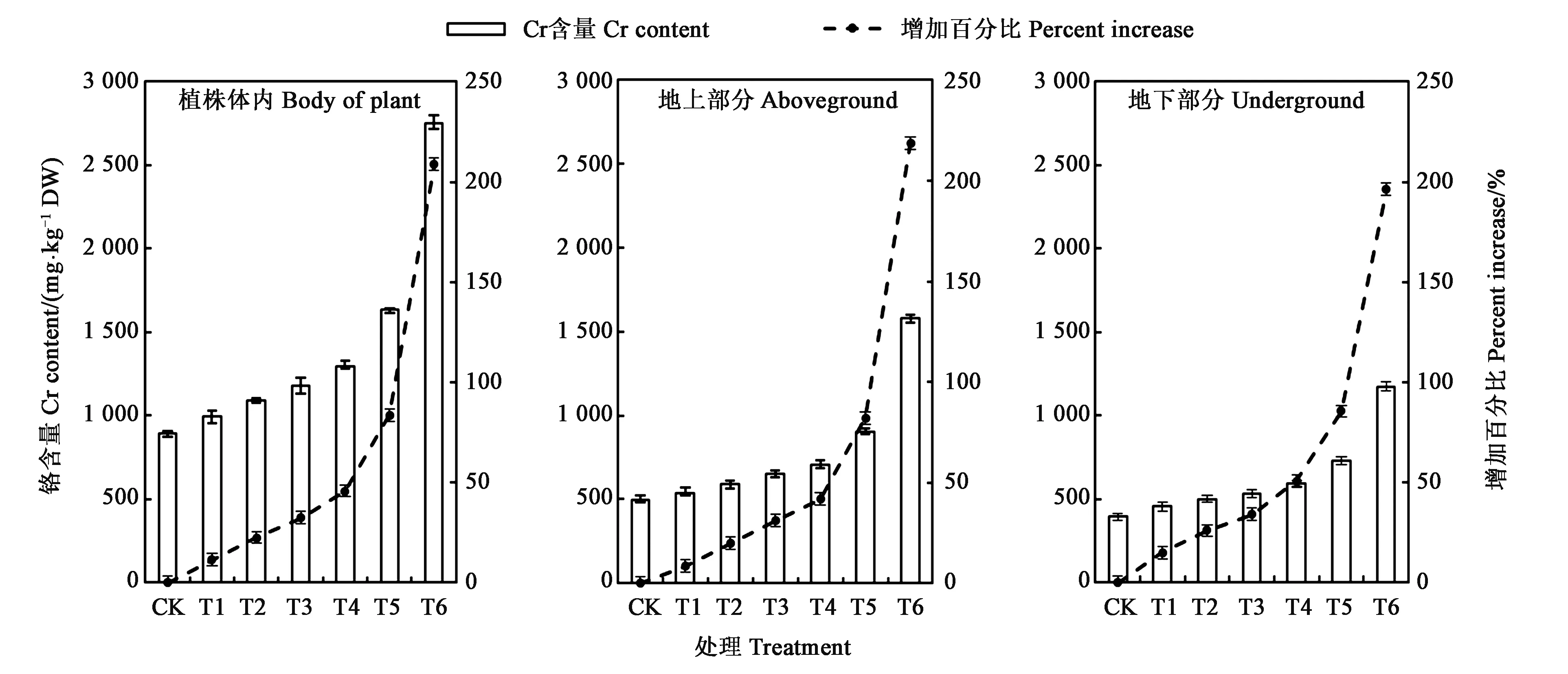
图1 不同处理下延胡索体内、地上部分、地下部分铬含量Fig.1 Chromium content in the body, aboveground and underground of Corydalis yanhusuo under different treatments
2.2 铬处理对延胡索株高和生物量的影响
图2为不同浓度Cr6+处理下延胡索植株株高与生物量的变化情况。随延胡索植株体内Cr6+含量的增加,延胡索植株株高、生物量总体呈现先增后减趋势。当土壤中Cr6+浓度低于T2时,Cr6+浓度越高,植株株高、生物量越大,且在T2时,与CK相比显著增加(P<0.05);当土壤中Cr6+浓度增加至T4(131.32 mg·kg-1)时,延胡索植株的株高、生物量有所下降,但仍然高于CK;当土壤中Cr6+浓度进一步增加至T5时,延胡索生长受到抑制,其株高、生物量均低于CK,且较T2时减少近1/3,说明此时延胡索植株生长受到严重危害,生理、生化过程受到一定的阻碍,生长发育停滞。
2.3 铬处理对延胡索MDA含量的影响
丙二醛(MDA)是膜脂过氧化物最终分解产物,其含量的高低可反映细胞膜脂过氧化的程度和植物对逆境反映的强弱[2]。图3显示了外源添加不同浓度Cr6+对延胡索植株内MDA含量的影响。分析发现,当土壤中Cr6+浓度低于T1时,与CK相比,MDA含量增加并不明显(P>0.05);而当土壤中Cr6+浓度在T2时,MDA含量较CK显著增加(P<0.05),为CK的1.4倍;当处理浓度达到最大时(T6),MDA含量达CK的6.9倍。表明土壤中Cr6+浓度越高,对植株产生的伤害越大。
2.4 铬处理对延胡索抗氧化酶活性的影响
图4显示了不同Cr6+处理下,延胡索抗氧化酶活性的变化。SOD作为植物体内清除ROS的第一个关键酶,可以将超氧阴离子自由基歧化为H2O2[13]。由图4可知,当土壤中Cr6+浓度在67.32~323.32 mg·kg-1范围内,SOD活性相对CK增加百分比随Cr6+浓度的增高而呈现下降趋势,当浓度达到323.32 mg·kg-1时,SOD活性达到最小,与CK的SOD活性相比下降达94.18%。通过SOD歧化产生的H2O2可以通过CAT与POD的降解来清除[13]。延胡索植株细胞中CAT活性随外源添加Cr6+浓度的增加而表现为先增后减的趋势。当土壤中Cr6+浓度低于131.32 mg·kg-1时,延胡索植株中CAT活性均高于CK;在浓度为99.32 mg·kg-1时,CAT活性较CK增加百分比达到峰值为21.32%;在131.32~323.32 mg·kg-1浓度范围内,CAT活性表现出减少趋势;当外源Cr6+处理浓度达到最高,CAT活性最小,较CK减小了81.50%。现有研究显示,严重的逆境也可以增强POD的活性,因为逆境的处理会使得植株产生更多的过氧化物[14],植株在逆境下受到的损伤程度越严重,其POD活性则会随之增高。本试验中,POD活性随土壤中Cr6+浓度增大而增加,当浓度为323.32 mg·kg-1时,POD活性较CK增加174.68%左右。
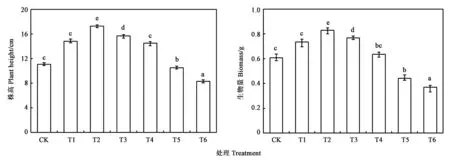
注:不同小写字母表示处理间差异在P<0.05水平具有统计学意义。Note: Different small letters indicate significant difference at P<0.05 level.图2 铬处理对延胡索株高和生物量的影响Fig.2 Effect of chromium treatments on plant height and biomass of Corydalis yanhusuo
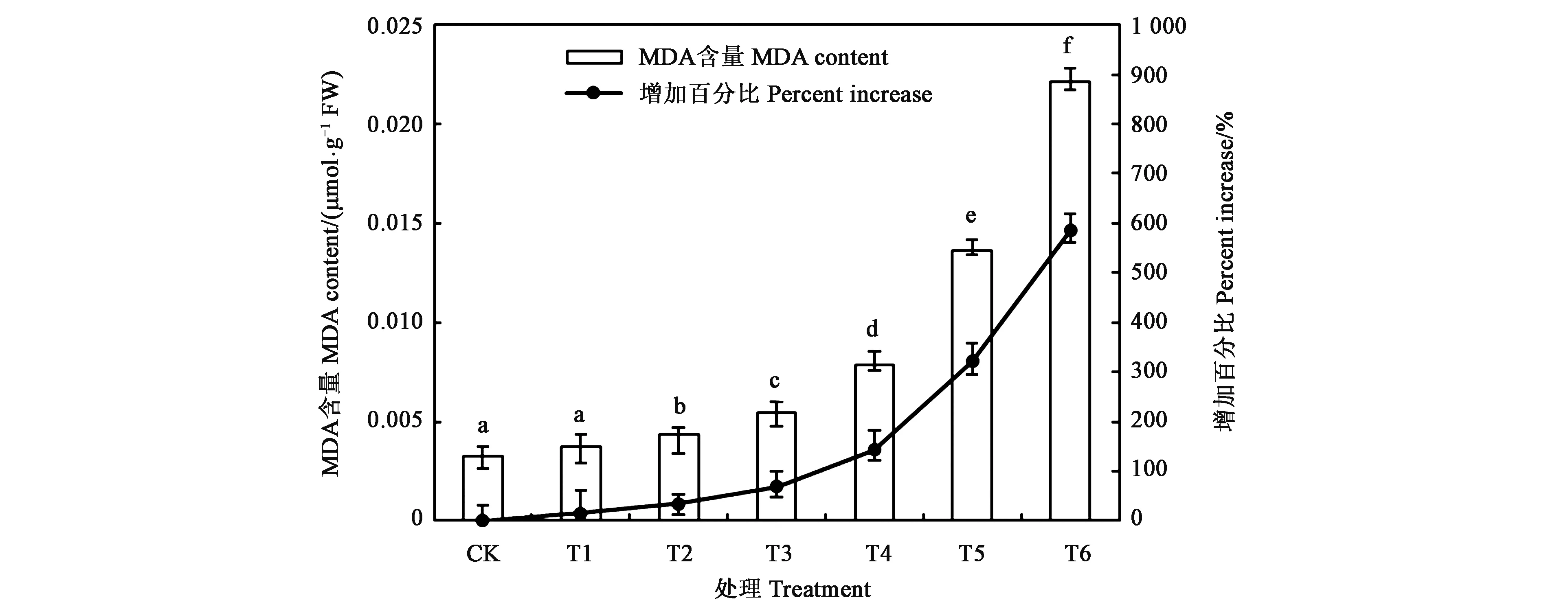
注:不同小写字母表示处理间差异在P<0.05水平具有统计学意义。Note: Different small letters indicate significant difference at P<0.05 level.图3 不同处理下延胡索MDA含量Fig.3 MDA content of Corydalis yanhusuo under different treatments
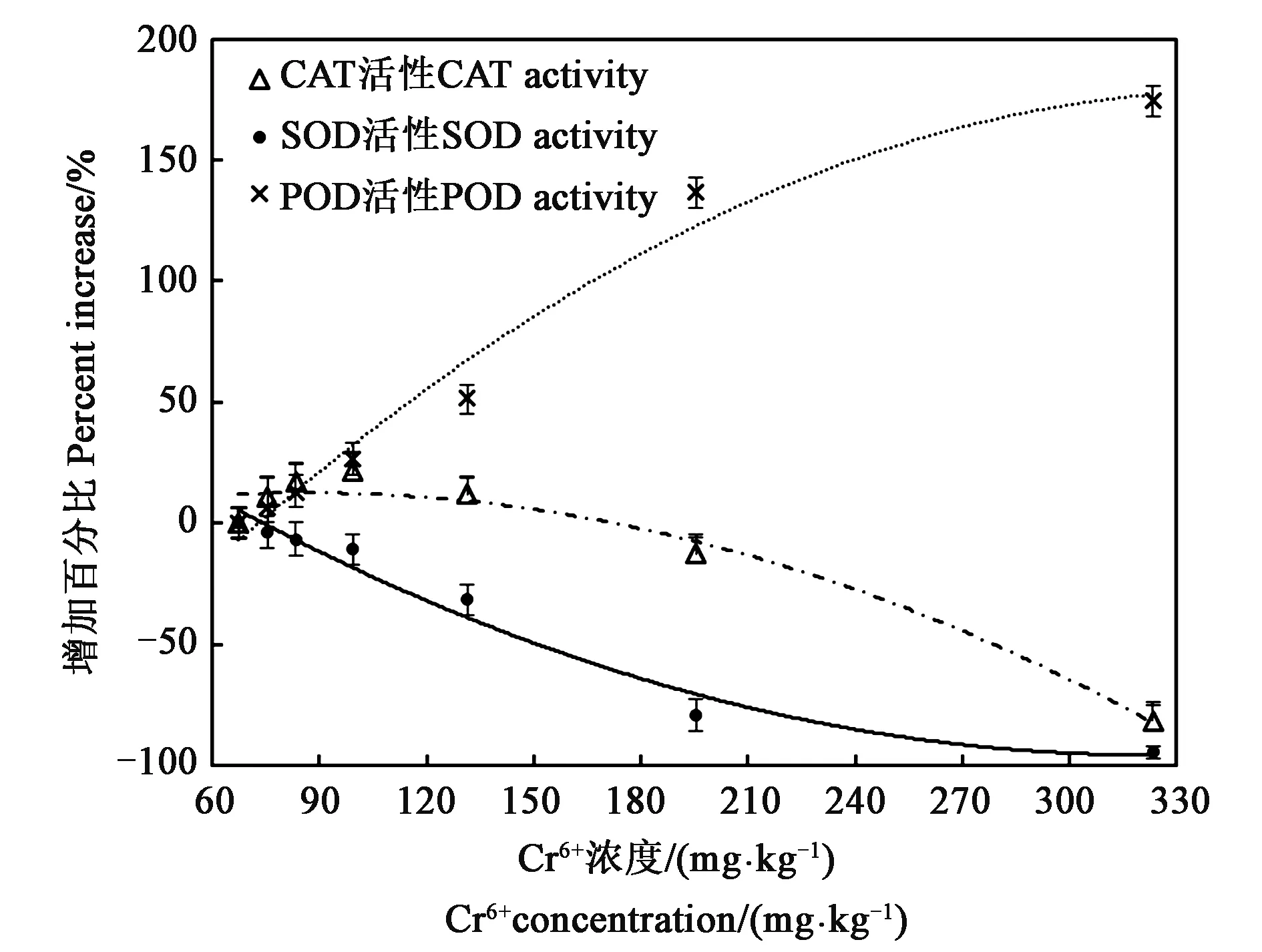
图4 不同浓度铬处理下延胡索SOD、POD、CAT活性增加百分比Fig.4 Percent increase of SOD, POD and CAT activities in Corydalis yanhusuo under different Cr6+ treatment
2.5 铬处理对延胡索GSH-AsA循环抗氧化物质的影响
正常情况下,GSH和AsA可以直接与超氧阴离子自由基等活性氧反应,也可通过GSH-AsA循环清除SOD歧化产生的H2O2[15]。从图5可以看出,在Cr6+处理下,随处理浓度升高,GSH、AsA含量均呈下降趋势,且AsA下降幅度高于GSH的下降幅度。当土壤中Cr6+浓度达到323.32 mg·kg-1时,GSH、AsA含量相对于CK分别下降61.28%和75.37%。
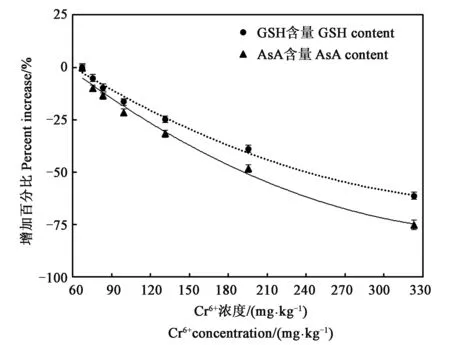
图5 不同浓度铬处理下延胡索GSH、AsA含量增加百分比Fig.5 Percent increase of GSH and AsA content in Corydalis yanhusuo under different Cr6+ treatment
3 讨论
3.1 延胡索对铬的吸收和转运
在相关研究中发现,一般植物对重金属的富集规律为地上部分重金属含量<地下部分重金属含量。Shanker等[16]研究表明,与地上部分相比,2个品种的大豆根系中都累积了大量的Cr,其种子中仅发现0.1%的Cr,而其根系之中存在的Cr高达98%。这与本研究中延胡索植株中重金属铬分布情况相反,这可能是由于延胡索根系吸收Cr6+后,Cr6+在其体内由地下部分向地上部分的迁移率较高,与此同时,Cr6+向上运输量大,从而导致茎叶中Cr6+含量较地下部分相对要高[17]。延胡索对Cr6+的这种转运机制使得其地下块茎药用部分的重金属积累更少,能够更好地被使用。
3.2 铬含量与延胡索生长的关系
重金属Cr是植物生长发育所必需的金属元素之一,但其含量超过植物体生长所需时,则会产生毒害作用,从而对植物细胞的结构和功能产生伤害[18]。黑淑梅[19]在关于Cr6+对小麦生长影响的研究中发现,当土壤中Cr6+浓度较低时,对玉米苗生长有明显的刺激作用;但浓度过高后,有明显的抑制作用。本研究结果表明,土壤中Cr6+浓度低于83.32 mg·kg-1时,延胡素植株株高、生物量均保持良好的增长趋势,这充分发挥了Cr作为必需元素的作用;随着土壤中Cr6+浓度的升高,对延胡素株高、生物量的影响逐渐显著,土壤Cr6+浓度为131.32 mg·kg-1的实验组,使得延胡索植株的株高、生物量显著减小,但仍高于CK;在此浓度之后则表现为延胡索株高、生物量均低于CK,证明在延胡索对Cr6+胁迫的耐受阈值为131.32 mg·kg-1。可能是因为土壤中高浓度的Cr6+胁迫对延胡索根系的细胞结构造成了破坏,使得延胡索体内Cr6+含量增加,从而导致机体的失衡。
3.3 铬对延胡索MDA含量和抗氧化系统的影响
MDA是植物体内ROS累积水平和细胞膜系统受损程度的评判指标[20]。在外源铬胁迫下MDA含量与处理浓度呈正相关,这说明Cr6+处理使得延胡索体内的ROS产生和清除平衡被打破,从而使得ROS大量积累,积累的ROS则引发了膜脂过氧化,因此MDA含量进一步增加,这与陈顺钰等[21]、高扬等[5]的研究结果一致。
Klaus等[2]与郭红叶[9]的研究都表明,植物抗氧化系统活性变化的特征是指示植物对环境胁迫响应类型的指标之一。本研究中,SOD活性降低,说明歧化超氧阴离子自由基的能力减弱,此时应该有大量ROS在植物体内积累,将导致植株生长受到抑制甚至死亡。但从试验中可知,在土壤中Cr6+浓度低于131.32 mg·kg-1时,Cr6+对延胡索的生长表现为促进作用。综合分析,出现这种现象可能由于SOD初始活性较高,因此,当土壤中Cr6+浓度低于131.32 mg·kg-1时,SOD仍然保持有较高的活性将超氧阴离子自由基歧化分解为H2O2,而POD与CAT活性也升高用以分解H2O2,因此,虽然此时植物体内产生的ROS增加,但利用抗氧化酶系统的清除能力,延胡索体内的ROS能保持在正常范围内,并且在此浓度下可能对延胡索体内生长基因的表达具有促进作用,所以延胡索植株在土壤中Cr6+浓度低于131.32 mg·kg-1时,株高、生物量均表现为增加趋势。但当土壤中Cr6+浓度达到195.32 mg·kg-1后,虽然POD活性有较大幅度的增加,但是由于MDA含量显著上升(P<0.05),且SOD对于超氧阴离子自由基的分解能力明显减弱,此时延胡索植株受到严重胁迫,因此其生长受到抑制。而GSH与AsA含量均表现出下降趋势,分析其主要原因,AsA的不断还原再生主要是GSH起作用,而GSH含量降低必然导致还原再生减弱,因此,AsA含量同样随之降低。本研究中用不同浓度Cr6+对延胡索植株进行处理的实验结果均与此相符,延胡索植株细胞内GSH含量与AsA含量呈现正相关关系,即伴随着GSH含量的减少AsA含量也随之减少。同时,延胡索植株内的Cr6+可诱导PCs的合成,而PCs的前体则为GSH,因此,当延胡索受到Cr6+的处理后,GSH用于合成PCs也将会使得GSH含量的减少。此时,GSH-AsA循环就受到限制,这导致H2O2清除同时也受到限制,因此导致H2O2在细胞中累积。同样,在土壤中Cr6+浓度低于131.32 mg·kg-1时,两者含量虽然减少,但其还是保持一定的清除能力以清除H2O2。综合以上抗氧化系统各指标的变化情况分析,在外源添加Cr6+浓度增加的情况下,包括SOD、GSH、AsA在内的大部分抗氧化酶活抗氧化剂,在外源添加Cr6+处理一开始时即表现为活性或含量降低,但在此种情况下,延胡索植株生长仍受到促进,分析认为,虽然受到了Cr6+的处理,但由于延胡索体内本身此类专性抗氧化酶与非专性抗氧化剂活性或含量较高,且一定浓度Cr6+的存在激发了延胡索体内其他酶类的表达,进而使得延胡索生长受到促进;但是一旦Cr6+浓度过高,抗氧化系统对植物的保护能力减弱,植物体内细胞结构以及生理生化反应遭受破坏,最终使得延胡索植株生长受到抑制[22]。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
舰船科学技术(2022年11期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
世界中医药(2020年5期)2020-04-19
保健与生活(2020年6期)2020-03-20
北方药学(2016年11期)2016-11-22
食品工业科技(2014年21期)2014-03-11
