
荜茇酰胺通过STAT3介导表观遗传调控下调胃癌细胞TERT表达的研究*
2020-02-27 05:19段超勤邹晓平陈志荣
胃肠病学 2020年8期
段超勤 闵 寒 邹晓平 陈志荣
南京医科大学附属苏州医院 苏州市立医院东区消化科1(215001)
南京大学医学院附属鼓楼医院消化科2
背景:荜茇酰胺(PL)是提取自中药荜茇的一种生物碱类化合物,许多研究表明,PL在体内、体外对多种肿瘤细胞具有抗肿瘤作用,但具体机制有待进一步探讨。目的:探讨PL对胃癌细胞TERT表达的调控机制。方法:分别以不同浓度PL、AG490处理胃癌细胞,CCK-8和平板集落形成实验检测胃癌细胞活性,实时荧光定量PCR检测TERT mRNA表达,蛋白质印迹法检测TERT、STAT3、p-STAT3、DNMT1蛋白表达,TRAP-ELISA法检测端粒酶活性,萤光素酶报告基因检测TERT基因启动子活性。结果:与对照组相比,PL组胃癌细胞活性明显下降,集落形成能力明显降低,TERT mRNA和蛋白表达、端粒酶活性明显下降,p-STAT3和DNMT1蛋白表达下降。AG490可明显抑制胃癌细胞活性以及p-STAT3、DNMT1、TERT蛋白表达。结论:PL可显著抑制胃癌细胞生长,其机制可能是通过STAT3介导的表观遗传调控TERT表达,有望成为治疗胃癌新的靶点药物。
胃癌是一种常见的恶性肿瘤,据最新统计,全球每年约有100万例新发胃癌病例和80万例胃癌相关死亡病例,是全球第五大最常诊断的癌症和第三大癌症死亡原因[1]。在我国,胃癌的总发病率和死亡率在各种恶性肿瘤中分别位居第二和第三位,且呈上升的趋势[2]。近年来,随着免疫治疗、分子靶向药物、新辅助化疗等治疗方法的不断进步和完善,胃癌治疗策略有所改善,胃癌患者的总体生存率有所提高,但进展期胃癌患者的预后仍不理想,5年生存率低于40%[3]。化疗仍是进展期胃癌患者最主要的治疗方法,但具有不良反应多、患者耐受性差、有效率低、并发症多等诸多问题[4]。因此,开发新的药物以改善进展期胃癌患者的生存预后是亟需解决的临床问题。荜茇酰胺(piperlongumine, PL)是从胡椒属植物荜茇中分离出来的一种天然植物碱,在体内外可有效地抗肿瘤细胞毒性[5]。本研究的前期研究发现PL对胃癌细胞的端粒酶逆转录酶(TERT)具有明显的抑制作用[6],但具体的调控机制不详[7]。本研究通过检测PL处理过的胃癌细胞中TERT、STAT3、DNA甲基转移酶1(DNMT1)表达,旨在探讨PL调控胃癌细胞TERT表达的机制。
材料与方法
一、细胞株、主要试剂
人胃癌细胞株AGS、HGC27购自中国医学科学院基础医学研究所细胞资源中心,由南京医科大学附属苏州医院肿瘤实验室冻存保管。TERT萤光素酶报告基因质粒由南京大学医学院附属鼓楼医院消化科友情提供,PL(Sigma-Aldrich Co. LLC.),酪氨酸激酶抑制剂AG490(Selleck Chemicals),CCK-8细胞增殖/细胞毒性检测试剂盒[东仁化学科技(上海)有限公司],TERT、信号转导与转录激活因子3(STAT3)抗体、磷酸化信号转导与转录激活因子3(p-STAT3)抗体、DNMT1抗体(Santa Cruz Biotechnology, Inc.),羊抗鼠、羊抗兔二抗(Cell Signaling Technology, Inc.),端粒酶活性检测试剂盒(美国Millipore),Lipofectamine 3000(美国Invitrogen公司)。
二、方法
1. 细胞培养:将胃癌AGS和HGC27细胞以含10%胎牛血清和1%双抗(100 U/mL青霉素+100 μg/mL链霉素)的RPMI 1640培养基培养于5% CO2、37 ℃恒温细胞培养箱内,常规消化传代培养。
2. 细胞处理:取对数生长期AGS和HGC27细胞,按后续实验需求以5×103/孔接种于96孔板(细胞活性检测),或以1 000/孔细胞接种于6孔板,12 h细胞贴壁后加入终浓度为0、5、7.5、10 μmol/L PL,每2~3天更换培养基1次(集落形成实验),或以2×105/孔接种于6孔板(除细胞活性和细胞集落形成实验外的其余检测),孵育24 h待细胞贴壁后进行相应处理。具体分组处理方式包括:①加入终浓度为0、7.5、10 μmol/L PL,孵育24 h;②加入终浓度为7.5 μmol/L PL,孵育12 h、24 h、36 h、48 h、60 h;③加入7.5 μmol/L PL孵育24 h,并设置不予任何处理的对照组;④将100 μmol/L AG490加入HGC27细胞,培养24 h。
3. 细胞活性检测(CCK-8法):AGS和HGC27细胞经相应处理后,弃培养基,加入含10% CCK-8的培养基避光孵育1.5 h,以酶标仪测定450 nm波长处的吸光度(A)值。细胞相对存活率=(A实验组-A空白组)/(A对照组-A空白组)×100%。
4. 集落形成实验:于6孔板中每孔接种1 000个胃癌细胞,过夜使其贴壁生长后,加入0、5、7.5、10 μmol/L PL培养14 d。待细胞长成肉眼可见的集落后,2 mL甲醇固定15 min,行0.5%结晶紫染色15 min,显微镜下统计每组集落形成个数。
5. 实时荧光定量PCR:不同浓度PL处理AGS和HGC27细胞后,以Trizol试剂提取细胞RNA,反转录为cDNA,行实时荧光定量PCR。TERT引物上游:5’-GCG TTT GGT GGA TGA TTT CT-3’,下游:5’-CAG GGC CTC GTC TTC TAC AG-3’;内参β-actin引物上游:5’-AGC GAG CAT CCC CCA AAG TT-3’,下游:5’-GGG CAC GAA GGC TCA TCA TT-3’。引物由生工生物工程(上海)股份有限公司合成。采用2-ΔΔCt法计算目的基因的相对表达情况。
6. 蛋白质印迹法:AGS和HGC27细胞经相应处理后,胰酶消化、收集细胞,预冷PBS洗涤2次,加入适当裂解液4 ℃或冰上裂解15 min,12 000 r/min离心15 min,BCA法蛋白定量,将各组蛋白调整为同一浓度,沸水煮5 min。取20~40 μg总蛋白上样,8%~12% SDS-PAGE电泳分离1~2 h;蛋白转移至PVDF膜,5%脱脂牛奶封闭2 h;加入1∶1 000~1∶2 000稀释的相应一抗,4 ℃孵育过夜,TBST洗涤3次;加入相应二抗孵育2 h,TBST洗涤3次;ECL显影,化学发光成像系统捕获并保存图像。
7. 端粒酶活性检测:采用TRAP-ELISA法检测端粒酶活性,以7.5 μmol/L PL处理HGC27细胞,分别在不同时间点按试剂说明书步骤进行细胞收集、处理,检测630 nm波长处的相应A值。
8. 启动子萤光素酶报告基因检测:待接种于6孔板的细胞密度生长至60%~70%时,将pGL3-SV40(阳性对照质粒)、pGL3-3778和pGL3-2089报告基因载体质粒(将TERT启动子近端的3778bp和2089bp片段亚克隆至pGL3载体构建而成),以脂质体-DNA复合物形式加入细胞培养孔,质粒转染按照Lipofectamine 3000说明书进行,24 h后更换培养基,加入7.5 μmol/L PL再培养24 h,收集并处理细胞后用双萤光素酶报告分析系统(Promega)进行检测分析。
三、统计学分析

结 果
一、PL抑制AGS和HGC27细胞增殖
以0、5、7.5、10 μmol/L PL处理AGS和HGC27细胞24 h后,CCK-8法检测结果显示,PL可抑制两株胃癌细胞增殖,且抑制作用呈剂量依赖性(图1A)。以相应浓度PL处理两株细胞2周,肿瘤细胞集落形成能力明显下降(图1B)。
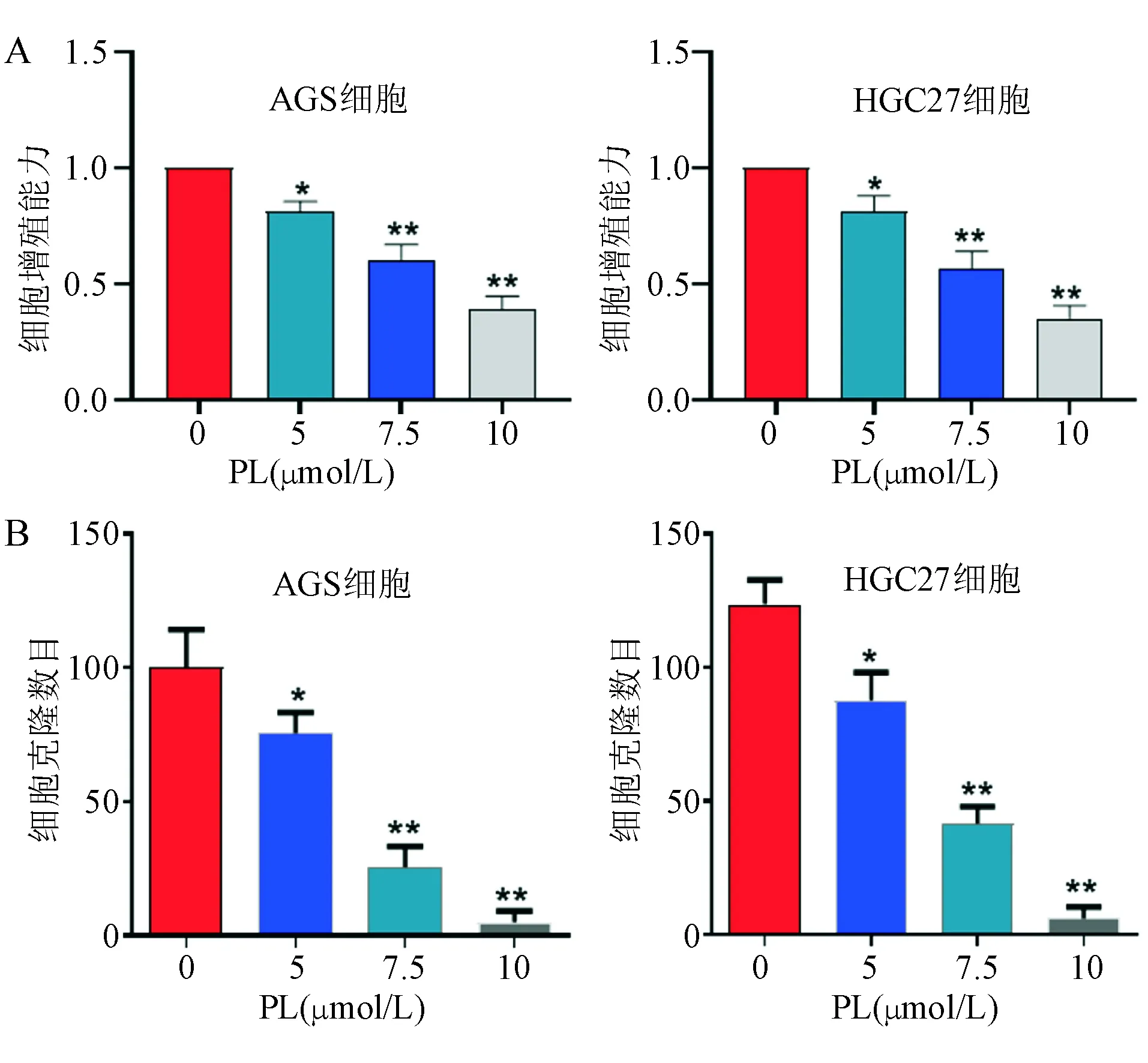
*P<0.05, **P<0.01
二、PL抑制胃癌细胞TERT基因转录和蛋白表达
实时荧光定量PCR和蛋白质印迹法结果显示,PL可显著降低AGS和HGC27细胞TERT mRNA和蛋白表达,且作用呈剂量依赖性(图2A-2B)。

1 Da=0.992 1 u
三、PL抑制胃癌细胞端粒酶活性
TRAP-ELISA法结果显示,7.5 μmol/L PL可呈时间依赖性地降低细胞端粒酶活性,与抑制HGC27细胞生长的趋势相一致(图3A-3B)。

*P<0.05, **P<0.01
四、PL通过STAT3介导甲基化抑制TERT表达
与对照组相比,7.5 μmol/L PL对TERT启动子萤光素酶活性无明显抑制作用(图4A)。
蛋白质印迹法结果显示,PL可明显抑制p-STAT3和DNMT1表达,且呈剂量依赖性(图4B)。给予AG490处理HGC27细胞48 h后,细胞活性受到显著抑制(图4C),p-STAT3、DNMT1、TERT蛋白表达明显降低(图4D)。
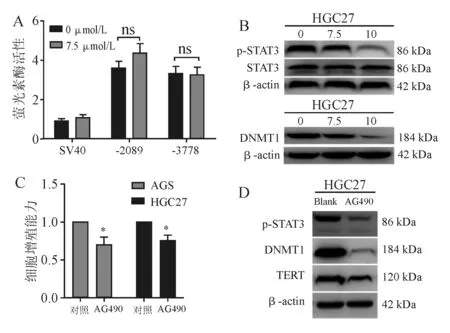
*P<0.05
讨 论
近年来,诸多体内、体外研究结果表明PL对多种肿瘤细胞具有明显的抗肿瘤作用,且未发现明显的毒副作用,其可能通过升高癌细胞内的活性氧水平、诱导氧化应激,从而促进细胞发生凋亡、阻滞细胞周期[8-10]。本研究发现,PL在体外对胃癌细胞具有显著的抗肿瘤作用,可抑制胃癌细胞活性、TERT mRNA和蛋白表达,并明显抑制胃癌细胞STAT3蛋白表达。
TERT是端粒酶的催化成分,是端粒酶活性的限速因子,目前发现超过90%的癌症中可见TERT表达,并在各种癌细胞的持续增殖中发挥重要作用[11]。目前认为TERT是可能肿瘤治疗的潜在靶点。有研究[12-14]显示小干扰RNA敲减TERT后可抑制癌细胞增殖,诱导细胞凋亡,并可增强化疗的敏感性。基于调控端粒酶的肿瘤治疗策略已成为数十年来的研究热点,并发现了一些潜在的端粒酶抑制剂如萝卜硫素等,但由于该过程的复杂性,目前仍未研发出理想的抑制剂应用于临床[15]。本研究发现,PL对TERT具有明显的抑制作用,有望成为新的TERT抑制剂。
表观遗传调控是指在不改变DNA序列的情况下发生的基因功能的可遗传变化,主要调控方式包括DNA甲基化、组蛋白乙酰化、组蛋白甲基化、非编码RNA调控等。表观遗传调控不仅在器官和个体发育中起重要作用,而且在许多疾病尤其是在肿瘤的发生、发展中起有重要作用[16-17]。DNA甲基转移酶在多种肿瘤细胞中高表达,与抑癌基因启动子甲基化密切相关,DNMT1可能在胃癌的发生、发展和治疗过程中起有重要作用[18-19]。本研究中,以PL处理后DNMT1表达下调,TERT表达相应下调,提示PL可能通过DNMT1影响TERT表达。
有研究报道,在恶性T淋巴细胞中,STAT3通过与DNMT1基因启动子中的STAT3 SIE/GAS结合位点结合调控DNMT1的转录[20]。STAT3在不同肿瘤细胞株中均可调控TERT表达[21-22]。AG490是一种可特异性抑制JAK/STAT3信号通路的酪氨酸激酶抑制剂,在胃癌和膀胱癌细胞中具有明显的抗肿瘤作用[23-24]。本研究以AG490处理胃癌HGC27细胞后,DNMT1和TERT表达均明显下调,提示在胃癌细胞中JAK/STAT3信号通路可能参与了PL介导的DNMT1下调,从而抑制TERT表达。
总之,PL具有显著的体外抗胃癌作用,其机制可能是通过JAK/STAT3途径下调DNMT1,进而抑制TERT表达。表观遗传调控在胃癌细胞PL-JAK/STAT3-DNMT1-TERT过程中起有重要作用,PL通过表观遗传调控影响TERT DNA甲基化,有望研发成为新的胃癌治疗药物。
猜你喜欢
生殖医学杂志(2022年10期)2022-10-19
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年5期)2021-07-22
西南林业大学学报(2021年3期)2021-05-03
中小学德育(2020年11期)2020-03-18
分析化学(2017年12期)2017-12-25
教育界·上旬(2016年12期)2017-05-25
标记免疫分析与临床(2016年9期)2016-11-21
腹腔镜外科杂志(2016年10期)2016-06-01
家庭医学(2015年6期)2015-07-03
