
连云港市枫树湾景区枫香种群结构与分布格局
2020-02-21 04:19:30李冬林江浩王火祝亚云万欣何冬梅
生态环境学报 2020年1期
李冬林,江浩,王火,祝亚云,万欣,何冬梅
江苏省林业科学研究院,江苏 南京 211153
种群是构成森林群落的基本单位,其结构不仅对群落结构有直接影响,并能客观地体现出群落的发展与演变趋势(Wu et al.,2002),在很大程度上反映种群与环境之间的相互关系(李先琨等,2002;Fulé et al.,1997)。种群的分布格局是由种群本身生物学特性、生境条件以及2者之间的相互作用所决定的,是对种群个体在水平空间上配置和分布状态做出的定量描述(Quinn et al.,1983;闫淑君等,2002)。研究种群分布格局可以为种群的水平结构进行定量描述,更重要的是可以揭示种群格局的成因,阐述种群分布与生境的关系,为预测群落的动态变化,制定种群的保护措施提供科学依据(Greig-Smith,1952)。
枫香(Liquidambar formosana)属金缕梅科枫香属,是我国南方常见的乡土阔叶树种。枫香具有喜阳、抗旱、耐瘠薄、生命力强的习性,在我国南方海拔1500 m以下的丘陵山区广泛分布,并成为我国北亚热带及部分暖温带森林的上层先锋树种(王发国等,2003;江聂等,2008;金保动等,2001)。枫香树型挺拔高耸,秋叶鲜红,在中、低海拔的丘陵岗地与常绿树种混生,形成红绿相映的多彩森林景观,为我国南方城市郊区重要的风景林资源。如我国湖南岳麓山(黄宗胜等,2005)、南京栖霞山(苏继申等,2003)和苏州天平山(周苏宁,1994)均是以枫香为主要观赏树种的风景名胜区,枫香占据森林植被的主林层。在当今森林旅游业和物流业快速发展的大背景下,城郊风景林正遭受生物入侵和人为活动的双重干扰,部分种群趋于衰退,并直接影响森林植被的林相特征和健康演替。因此,开展城市风景林资源的调查,分析种群动态和森林更新特点,并依此来制定种群的保护措施,促进风景林资源的结构优化和健康延续具有十分重要的理论和实践意义。目前,枫香的研究文献多涉及南方森林。张志华等(2012)研究了我国亚热带天目山自然保护区枫香种群的结构特征和空间分布格局;王丹等(2017)研究了地处我国西南的贵阳观山湖岸枫香种群的空间格局与环境因素的关系;邓送求等(2010)以南京紫金山为例研究了我国北亚热带落叶阔叶林中枫香种群的结构调整和演替规律,而对于地处北方暖温带,并以枫香为建群种的森林结构与动态规律的研究尚未见报道。本文通过样方法对江苏连云港枫树湾景区的枫香种群进行了系统调研,分析了群落内枫香种群的结构及分布格局特征,以期为枫树湾景区枫香种群的结构优化和林相维护提供一定的参考依据。
1 研究区概况
调查区位于江苏省连云港市连云区宿城街道枫树湾景区。区域坐标为 34°42′10″—31°39′36″N,119″25′50″—119″25′60″E。属典型的暖温带季风气候,常年平均气温14 ℃,1月平均温度-0.4 ℃,极端低温-19.5 ℃:7月平均温度26.5 ℃,极端高温39.9 ℃。年平均降水量920 mm,常年无霜期220 d。地带性土壤为片岩或片麻岩发育起来的灰棕色砾质棕壤,呈微酸性反应。
枫树湾森林植被保存完好,林相整齐。群落类型为典型的落叶阔叶林,上层主要组成树种为枫香,还夹杂有少量的朴树(Celtis sinensis)、糙叶树(Aphananthe aspera)、板栗(Castanea mollissima)、楸树(Catalpa bungei)、黄檀(Dalbergia hupeana)、野山楂(Crataegus cuneata)等。上层林分郁闭良好,平均郁闭度 0.85。下木和灌木层常见种类有茶条槭(Acer ginnala)、山胡椒(Lindera glauca)、菝葜(Smilax china)、野蔷薇(Rosa multiflora)、短柄枹(Quercus glanduliferavar.brevipetiolata)、木莓(Rubus swinhoei)、白檀(Symplocos paniculata)、猫乳(Rhamnella franguloides)、柘树(Cudrania tricuspidata)、野珠兰(Stephanandra chinensis)、美丽胡枝子(Lespedeza formosa)、蓬蘽(Rubus hirsutus)等。林下野生草本植物生长茂密,主要有络石(Trachelospermum jasminoides)、荩草(Arthraxon hispidus)、海金沙(Lygodium japonicum)、麦冬(Ophiopogon japonicus)、蕨(Pteridum aquillinum)、透骨草(Phryma leptostachya)、荠蒿(Artemisia anomala)、紫露草(Tradescantia ohiensis)、雀麦(Bromus japonicus)等。
2 研究方法
2.1 样地调查
在前期全面踏查的基础上,于2018年7—8月选取集中连片、群落完整、有代表性的地段设置调查样地7个(编号 Q1、Q2、Q3、Q4、Q5、Q6、Q7),单个样地面积为20 m×20 m,各个样地的生境特征见表 1。采用相邻格子法(罗文等,2010)将各样地分成5 m×5 m的小样方16个,并分别进行群落学调查。分层调查每个小样方内所有乔木树种的胸径(DBH)、树高、冠幅和灌木及下木的种类、株数及盖度,同时记录所有样方内出现的草本植物种类、盖度及多度情况。
群落调查指标的计算:乔木重要值=(相对密度+相对频度+相对显著度)/3;草灌木重要值=(相对盖度+相对高度)/2(宋永昌,2001)。
2.2 种群结构分析
由于枫香为阔叶大乔木,个体生长迅速、材质坚硬,种群数量多,用生长锥钻取木芯较为困难,种群个体的年龄结构不易测定,故采取空间代替时间的方法(康华靖等,2007;吴承祯等,2000),即用调查立木的大小级代替年龄结构来分析种群的结构和动态变化。参考有关文献,并结合枫香的自然生长进程,本研究采用以下标准进行立木大小级的划分(上线排外):Ⅰ级幼苗DBH<2.50 cm,Ⅱ级DBH 2.5—7.5 cm;Ⅲ级DBH 7.5—12.5 cm;Ⅳ级DBH 12.5—17.5 cm;Ⅴ级DBH 17.5—22.5 cm。Ⅵ级DBH 22.5—27.5 cm;Ⅶ级DBH 27.5—32.5 cm;Ⅷ级DBH 32.5—37.5 cm;Ⅸ级DBH 37.5—42.5 cm;Ⅹ级DBH 42.5—47.5 cm;Ⅺ级DBH 47.5—52.5 cm;Ⅻ级DBH>52.5 cm。以野外样方调查的数据统计各大小级的株数,以大小级比率为横坐标,大小级为纵坐标,绘制枫香种群的年龄结构。
2.3 种群存活曲线与静态生命表的编制
以各大小级的自然对数(ln)为横坐标,以现存个体数量的自然对数(ln)为纵坐标绘制枫香种群的存活曲线(金则新,2001)。统计各个大小级内枫香株数,依据野外调查的各龄级的存活数(ax)及标准存活量(lx),计算出各龄级的死亡数(dx)、死亡率(qx)、平均存活数(Lx)、从第x径级起超过x径级的有活个体总数(Tx)、生命期望(ex)等指标,完成种群静态生命表的编制(姜在民等,2018)。
2.4 种群分布格局分析
采用聚集度指标进行种群空间分布格局的分析(范繁荣等,2008;康佳鹏等,2019)。统计指标包括:扩散系数(C)、负二项参数(K)、扩散型指数(Iδ)、Cassie指标(Ca)、丛生指标(I)、平均拥挤指数(m*)、聚块性指标(m*/m)。
(1)扩散系数(C)


表1 枫树湾枫香种群不同样地的生境特征Table 1 Habitat condition of different sampling plots of Liquidambar formosana population in MapleTree Bay
扩散系数法C是检验种群是否偏离随机性的一个系数。若C=1,则种群随机分布;若C>1,则为集群分布;若C<1,则为均匀分布。显著性检验采用t检验。t值通过以下公式计算,然后t值与t0.05(n-1)比较定其差异显著性。

(2)负二项参数(K)

当K<0时,种群为均匀分布;当K>0时;种群为集群分布;当K>8时,种群为随机分布。
(3)扩散型指数(Iδ)

式中,n为样方数目,N总株数,xi为每样方内株数,fi为株数的频率。如果Iδ=1,表明个体是均匀分布的;如果Iδ<1,趋向于随机分布;若Iδ>1,则为集群分布。
(4)Cassie指标(Ca)

式中,S2为取样单位的方差;为每个样方中个体的平均数目。当Ca=0时,种群为随机分布;当Ca>0时,种群为集群分布;当Ca<0时,种群为均匀分布。

(5)丛生指标(I)当I<0时,为均匀分布;当I=0时,随机分布;当I>0时,为集群分布。
(6)平均拥挤指数(m*)

平均拥挤度(m*)反映了样方内生物个体的拥挤程度。当m*>时,为集群分布;当m*=时,为随机分布;当m*<时,为均匀分布。
(7)聚块性指标(m*/m)

当m*/m<1时,为均匀分布;当m*/m>1时,为集群分布;当m*/m=1时,为随机分布。
3 结果与分析
3.1 枫香群落的结构特征
连云港枫树湾森林群落为典型的落叶阔叶林,表2为不同样地群落中乔木层物种的重要值。可以看出,群落中枫香的优势地位比较明显,7个样地的合计重要值为561.477%,远远大于所有样地中其余树种的重要值;次优势种为朴树,其合计重要值为35.279%;糙叶树的合计重要值为25.157%,为第 3优势种。其它伴生的乔木种类的还有楸树(13.594%)、茶条槭(15.910%)、野山楂(10.124%)、板栗(3.042%)等,但重要值普遍较小。由此可知,枫香在群落中占绝对优势,呈单一优势群落。
3.2 枫香种群的龄级结构
种群年龄结构分析是探索种群动态有效方法,是揭示种群结构现状和更新策略的重要途径之一(梁士楚等,2002)。从图1可以看出,枫香种群的年龄结构呈现一定程度的偏态分布,但不同样地之间的年龄结构存在一定差异。样地 Q1中枫香种群的年龄结构呈现明显的偏峰变化,即Ⅰ、Ⅱ级幼苗、幼树的数量较多,两者大小级合计占比为70.69%,表明优苗幼树数量多,群落更新良好,但Ⅲ级以上成年大树数量较少,大小级合计占比仅为29.31%,样地Q2也呈现类似的变化。Q3、Q4、Q5、Q74样地的年龄结构基本一致,呈现幼龄级幼苗偏少、中龄级幼苗偏多的仿锤形变化,即幼龄级(Ⅰ级和Ⅱ级)的幼树幼苗比例不足,甚至缺失(Q3缺少Ⅱ级、Q4和Q5缺少Ⅰ级),但中龄级(Ⅲ级和Ⅳ级)及以上个体数量较多,如样地Q3中Ⅲ级木占22.22%,Ⅳ级占44.44%,样地Q4中Ⅳ级木占45.45%,Q5中Ⅶ级木占38.46%。样地Q6有一定的幼苗幼树,但存在一定的间断,个别龄级缺乏,表现为种群结构的完整性不良。不同样地间枫香种群的径级结构所表现出的这种异质性特征应该与不同样地所处的立地生境及人为干扰活动素有关。
从调查结果来看,第Ⅶ级以上种群个体分布普遍较少,说明枫树湾枫香林普遍存在的是中幼龄林木。但值得注意的是个别样地出现了Ⅺ以上的大树,如样地Q6中出现了胸径56.10 cm和64.5 cm的大树2株(Ⅻ级),样地Q7出现51.50 cm大树1株(Ⅺ级),并占据主林层。所以,尽管少量样地中种群幼年小树缺少或不足,但少量大规格树木的存在提高了重要值,枫香树在该种群中优势地位仍然明显(表2)。

表2 不同样地枫香群落乔木层物种重要值Table 2 The importance values of the arborous layer species in different plots of Liquidambar formosana
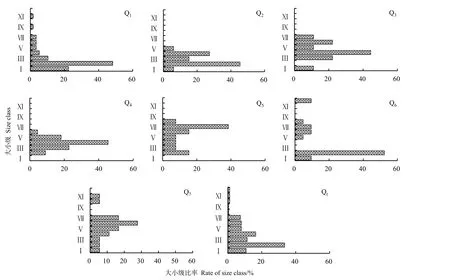
图1 不同样地枫香种群大小级结构Fig. 1 Size structure of Liquidambar formosana population in different plots
3.3 枫香种群的存活曲线
存活曲线是一条反映种群个体在各年龄的存活状况的曲线,是种群统计中最常见和最直观的方法(赵学农等,1996)。通过分析存活曲线,可以深刻了解树木种群年龄结构的动态变化及其内在演替规律。图2为调查区不同样方枫香种群的存活曲线。按E. S. Deevey的划分(Deevey,1947),存活曲线大致有3种基本类型,DeeveyⅠ型是凸曲线,该型的种群绝大多数都能存活到该物种年龄,早期死亡率较低。但当存活到一定生理年龄时,短期内几乎全部死亡,种群趋于衰退;DeeveyⅡ型是直线型,也称对角线型,属于该型的种群各年龄的死亡率基本相同,种群趋于理想性稳定,故又名稳定型种群;Deevey Ⅲ型是凹曲线,早期死亡率高,一旦存活到某一年龄段,死亡率降低,种群趋于稳定(姜在民等,2018)。
为了更准确地确定各样地存活曲线的特征,将各样地的存活曲线进行线性回归,配合直线回归方程并检验其显著程度(表3)。由表3可知,除样地Q1外,其它枫香种群的存活曲线的线性关系均不显著(P>0.05)。由此可以推断,样地Q1中枫香种群整体存活曲线为显著的直线型,即稳定型(DeeveyⅡ),种群中有足够数量的幼苗幼树及少量大树,存活曲线略有凸起更接近于直线,大小级结构趋于金字塔形(图 1)。其余样地种群的存活曲线呈现不规则。而对比各样地的存活曲线,样地Q3、Q4、Q5中种群存在着不同程度的个体缺失,中树、大树所占比例较多,幼苗、幼树数量普遍较少,后备更新资源严重不足,不利于种群的世代延续,大致上符合凸型曲线(DeeveyⅠ)型变化,总体上属于衰退型种群。比较而言,样地Q2幼龄期的幼苗幼树数量多,死亡率较低,拥有较多的后备更新资源,而中龄级及以上幼树个体死亡率偏高,大规格成年个体日渐稀少,并出现缺失,基本上仍符合凸型曲线(DeeveyⅠ)的种群变化特征。样地Q6、Q7的存活曲线几乎呈断点凸型,种群中有一定的幼树幼苗数量,也有一定数量的中树和大树,但在大龄级某一区段则出现断点,种群整体不稳定,不利于种群的世代延续。

图2 不同样地枫香种群存活曲线Fig. 2 Survivor curve of Liquidambar formosana population in different plots

表3 不同样地枫香种群存活曲线的直线回归方程Table 3 Linear regression equation of the survival curves of Liquidambar formosana population in different plots
3.4 枫香种群的静态生命表
借助于静态生命表可以直观地比较不同年龄段个体的数量变化,进而推断种群的动态变化趋势(魏新增等,2008)。静态生命表常采用同一时间(或某个调查期)内收集到的某一种群所有个体的大小结构编制而成,它反映了多个世代重叠的年龄动态历程中的一个特定时间,而不是对同生种群的全部生活史追踪(蔡小英等,2006)。枫树湾枫香种群的静态生命表见表 4。可见,种群第Ⅰ、Ⅲ和Ⅹ龄级的死亡率(qx)为负值,而第Ⅶ龄级的qx最大(85%),说明枫香幼龄期幼苗幼树比较适合于林下的自然生境,个体之间竞争较弱,出苗后一般死亡的可能性小。但到了大龄级以后,林内个体之间出现了激烈竞争,侧方庇荫增加,对于强阳性的枫香带来一定的威胁,加上生理衰老作用会造成一定数量的个体消亡,种群数量日渐减少。个体存活率(ax)具有波动性,在Ⅰ龄级后总体上随径级的增大而减少,其中幼苗阶段个体数量最多。期望寿命(ex)表达的是个体的平均生存能力(姜在民等,2018;蔡小英等,2006)。随着龄级的增加,ex出现了多次的波动,幼苗与壮龄阶段的生命期望值低于幼、中树阶段,Ⅲ龄级幼树出现拐点(3.67)。由于大龄级个体的跨越式分布,导致Ⅷ级以上个体局部年龄段的ex估计过高(表4)。
3.5 枫香种群的空间分布格局
种群是物种在特定自然条件下的生存方式,其现存的分布格局可从侧面反映种群的对环境的利用程度,并对种群的延续和发展带来深远的影响(魏新增等,2008;张文辉等,2005)。对枫香种群分布格局指标的统计计算得知,扩散系数(C)为4.8750—12.5989之间,均大于1(表5)。查t值表,t15,0.05=2.131,因此7个样地中所有t值均大于t15,0.05,则S2/对1的偏离显著,说明枫树湾景区的枫香种群分布格局均为集群分布(Clump)。其它分布格局的指标,如负二项参数(K)、扩散型指数(Iδ)、Cassie指标(Ca)、丛生指标(I)等也都呈现集群分布。比较不同样地的枫香种群的空间分布,不同样地的测定指标差异较大,说明集聚程度存在一定的差异。就拥挤指数(m*)来看,Q1、Q2样地的m*明显要大于其它样地,表现为种群内立木株数较多(分别为58和118株),个体规格偏小,个体竞争激烈;而两样地扩散系数(C)也明显较高,已经远远偏离了随机性分布。而样地Q6、Q7的生境偏僻,枫香立木个体规格较大,个体之间的竞争已趋于稳定,其聚集程度趋于弱化,其C值和m*明显小于其它样地。

表4 枫树湾枫香种群静态生命表Table 4 Static life table of Liquidambar formosana population in Maple Tree Bay
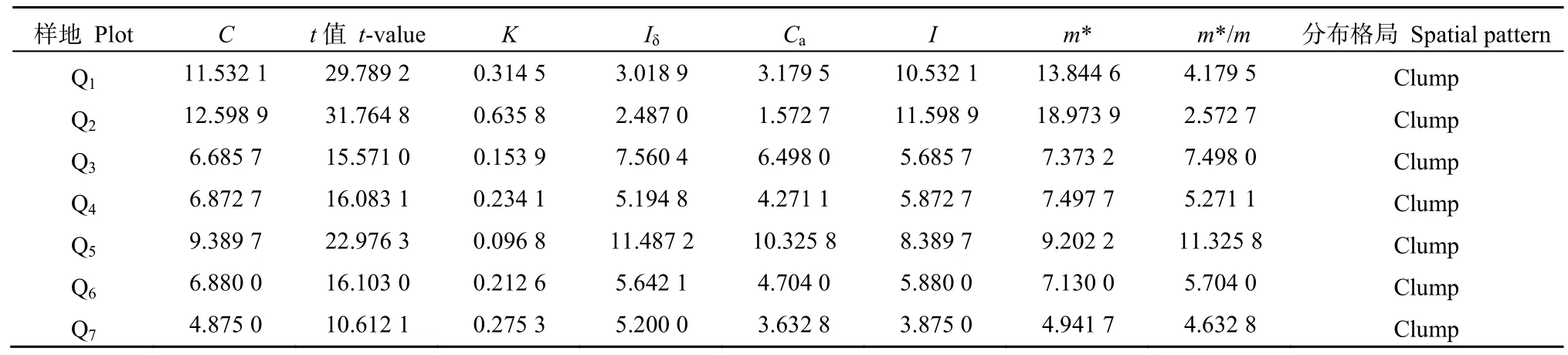
表5 枫树湾枫香种群空间分布格局Table 5 The spatial distribution pattern of Liquidambar formosana population in Maple Tree Bay
为了比较上述7个指标间的相互关系,用相关分析检验它们之间的相关性见表6。从表6可看出,C、I、m*之间,以及Ca与Iδ、m*/m均呈极显著的正相关(P<0.01),且相关系数几乎等于1,说明它们表征的生态学意义基本相同。K与Iδ、Ca、m*/m之间呈显著的负相关(P<0.05),说明它们表征的生态学意义相反。
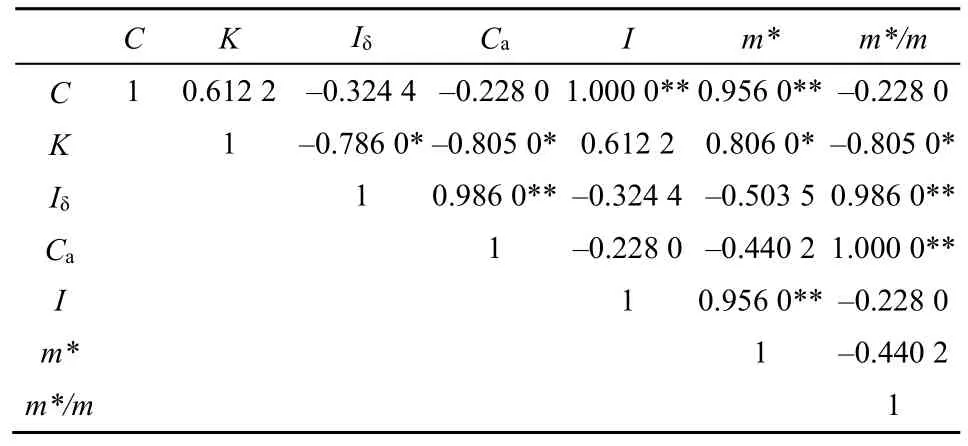
表6 各聚集指数之间的相关矩阵Table 6 The correlation coecients matrix of C, K, Iδ, Ca, I, m* and m*/m
4 讨论
(1)枫香起源古老,是地球上第三纪孑遗植物(王发国等,2003),喜温暖湿热的气候特征,在中国广泛分布于秦岭淮河以南的亚热带地区,此线以北一般只有零星栽培(周光裕,1995)。而在我国东部沿海、纬度偏北、冬季寒冷的江苏连云港保存有相当面积的天然枫香种群实为罕见,并成为我国枫香种群天然分布的北界。这一方面源于江苏连云港地处暖温带的南缘,东部临海,具有独特的海洋性气候特征,四季分明,光照充足,雨量适中。另外,枫树湾所在地宿城地处连云港市连云区东南部,三面环山,背风向阳,丰沛的水热和充足的阳光为枫香的栖居创造了优越条件,这是该区存在枫香野生种群的最根本的原因。值得注意的,本调查中还发现个别样地出现了Ⅶ以上的大规格树木(如胸径为64.5 cm个体1株,56.1 cm个体1株)。尽管这些大规格个体依据国际标准还称不上是古树名木,但是历史上云台山历经多年战乱,如此规格的枫香在此能得以保存实属不易。这些大规格树木资源是连云港极其宝贵的生物资源,具有特别重要的保存价值,一方面对于研究连云港市植物区系特征具有科学价值,另一方面对于维持枫树湾的森林风貌和红叶景观都具有重要意义,应重点予以保护。
(2)植物种群结构特征不仅能够体现种群内部个体的发展过程,而且也是对立地条件优劣及植物自身对环境适应性的反映(姜在民等,2018),其径级结构能很好地反应种群动态变化(Arista,1995)。从本研究的结果来看,枫香种群的年龄结构整体上呈现一定程度的偏态分布,并趋于衰退(DeeveyⅠ型),表现为幼苗和大树比例偏少,而且大小级结构存在着不同程度的个体缺失,多数样地的存活曲线呈现断点,种群后期延续性更新不良。其主要原因可能是森林上层林冠郁闭,林内光照较弱,但灌木生长茂密,占据了较多的下层空间,在一定程度上限制了枫香种子萌发和幼苗幼树的存活。另外,近年旅游业的快速发展也对地处城市郊区的枫香风景林构成了强烈干扰,加剧了枫香种群的衰退型演替。
(3)种群的水平空间分布格局是物种与环境长期相互适应、相互作用的结果,与自然生境及其所处群落中其余物种的竞争入侵等生态因子密切相关(苏日古嘎等,2009)。本研究表明,枫香种群分布格局表现为集群分布,这与王丹等(2017)、张志华等(2012)的研究结果相一致。枫香种群这种分布特征是其自身生物学特性和所处生境条件相互作用的必然结果。枫香的果实为圆球形蒴果,成熟后开裂,种子有窄翅,主要靠风力进行传播,在高山多风条件下散播飘逸带有很大的随机性。但枫树湾所处海拔偏低,三面环山,背风向阳,独特的小地形决定了枫香种群成熟种子的传播距离不会太远,分布格局总体上趋于集群分布。但是,枫树湾枫香种群在不同样地间的集群程度仍然存在一定差异。Q1、Q2样地海拔较低,坡度为0,地表土壤深厚,给林下种子的发芽创造了优越条件,尽管种内个体竞争激烈,但幼苗幼树的保存率依然较高,种群呈现的聚集程度最强;而样地Q6、Q7生境偏僻,海拔偏高,地表土壤浅薄,并有岩石裸露,尽管上层母树结实量大,但林下种子的天然发芽较为困难,自然更新不良,种群分布的聚集程度趋于弱化。因此,生境和自身生物学特性的双重作用导致枫树湾枫香种群的集群分布的复杂性。
(4)枫香是我国自然风景林资源最主要的组成树种,而枫树湾又是江苏连云港市著名的秋季赏叶地。枫香种群的结构维持和健康演替对维持连云港市宿城森林群落的林相外貌及可赏性,确保景区的可持续发展至关重要。采取适度的森林抚育手段,促进天然种群结构调整和群落稳定是当前生态风景林日常维护的重要措施。从枫树湾枫香种群现状来看,样地Q1、Q2种群内幼龄个体株数较多,竞争较为激烈,已出现明显的自然整枝和偏冠,不利于种群个体的树型发育和林相外貌,建议进行适度疏伐,减少侧方庇荫,促进幼树的健康成长;而生境偏僻的Q6、Q7样地,上层枫香个体规格较大,个体竞争已趋于稳定,但下层灌木植被丰富,林内光照不足,以及部分灌木植物的恶性竞争导致枫香天然种实发芽率低、幼苗幼树数量偏低,可适度进行人工抚育作业,适量清理杂灌,并开展枫香人工植苗或辅助更新,将有助于枫树湾景区枫香种群的结构优化和健康演替。
5 结论
连云港枫树湾森林群落为典型的落叶阔叶林,群落中枫香占绝对优势,呈单一优势群落。枫香种群的龄级结构不完整,年龄结构呈现一定程度的偏态分布,部分样地的存活曲线呈现断点,种群后期延续性更新不良,为衰退型种群。种群的分布格局呈集群分布,但是由于生境的影响不同样地间的集群程度存在一定差异。过分密集的样地种内竞争激烈,建议进行适度疏伐,增进林内光照;而生境偏僻、林下灌草过于繁茂的地段,建议进行适量清杂,促进种群的结构调整和健康演替。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
乡村地理(2022年2期)2022-05-30 10:48:04
环境与生活(2021年5期)2021-06-24 18:09:52
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
绿色科技(2016年23期)2017-03-15 15:41:56
科教导刊·电子版(2016年25期)2016-11-16 10:24:35
江苏农业科学(2016年5期)2016-07-23 15:59:31
当代贵州(2016年23期)2016-07-21 08:55:34
广西林业科学(2016年4期)2016-03-16 05:44:59
