
kshv-mir-k12-1-5p靶向调控CDKN1A影响卡波西肉瘤的细胞周期
2020-01-01 09:07:22彭靖淇吴秀娟普雄明
安徽医科大学学报 2019年12期
张 静,彭靖淇,吴秀娟,普雄明
卡波西肉瘤(Kaposi′s sarcoma,KS)是内皮细胞起源的恶性肿瘤[1]。卡波西肉瘤相关病毒(KSHV)属于γ2疱疹病毒[2],是KS最重要的病原体。KSHV主要通过编码miRNAs在KS中发挥作用[3]。miRNAs是非编码小分子RNA[4]。miRNAs通过结合靶基因3′UTR区下调或沉默靶基因的表达[5]。miRNAs调控超过1/3的人类基因[6],通过复杂的机制在各种细胞过程中发挥重要作用, 广泛参与肿瘤的形成和发展[7-8]。Wu et al[9]研究显示 :13个KSHV miRNAs在KS中表达全部上调,其中kshv-mir-k12-1-5p上调最高,KSHV miRNAs主要通过调控或改变细胞周期导致KS的发生[10],故推测kshv-mir-k12-1-5p可能与细胞周期的调控密切相关。细胞周期蛋白依赖性激酶抑制因子1A(CDKN1A)是功能强大的细胞周期负调控因子。miRNAs通过抑制靶基因的翻译而发挥其生物学功能[11]。现研究kshv-mir-k12-1-5p对靶基因(CDKN1A)的抑制作用及对KS细胞周期的影响,为KS的靶向治疗提供理论依据。
1 材料与方法
1.1 细胞株和主要试剂卡波西肉瘤细胞株(KS cell line, SLK)(美国NIH AIDS Research and Reagent Program提供); 293T cells(美国Open Biosystems公司);双荧光素酶报告系统试剂盒(美国Promega公司);Lipofectamine 2000、TRIzol试剂盒(美国Invitrogen公司);kshv-mir-k12-1-5p 模拟物(mimics)、kshv-mir-k12-1-5p抑制物(inhibitor)、mimics阴性对照(NC)、inhibitor NC(上海吉玛技术有限公司);RIPA裂解液、细胞周期分析试剂盒(上海碧云天生物技术公司);CDKN1A、GAPDH、HRP(美国Abcam公司)。
1.2 方法
1.2.1细胞培养和转染 使用对数生长期细胞密度为80%的SLK细胞(KS细胞)。将SLK细胞按每孔2×105细胞接种于96孔板中,培养24 h。Western blot、qRT-PCR及细胞周期PI实验均分5组进行:mimics组、inhibitor组、mimics NC组、inhibitor NC组、正常细胞组(normal cell group)。使用Lipofectamine 2000转染试剂将实验5组分别转染到SLK中,转染时间为48 h。
1.2.2双荧光素酶实验 将野生型(WT)和突变型(MUT) CDKN1A 3′UTR扩增并构建为pYr-mirTarget重组质粒。然后用Lipofectamine 2000转染试剂将重组质粒(1 μg)和kshv-mir-k12-1-5p mimics、mimics NC(50 nmol/L) 共转染293T细胞(5×104)。转染24 h后,使用双荧光素酶测定系统检测荧光素酶相对活性。
1.2.3Western blot(WB)检测 使用400 μl RIPA裂解液(50 mmol/L Tris/pH7.4、150 mmol/L NaCl、1% NP-40、0.5% sodium deoxycholate、0.1% SDS、sodium orthova-nadate、sodium fluoride、EDTA、leupeptin)提取细胞总蛋白。根据蛋白质含量,绘制标准曲线。制备层叠凝胶(5%)和分离凝胶(12%)。然后进行电泳(1.5 h、100 V)。当溴酚蓝到达凝胶板底部时,电泳结束。将蛋白转移到膜上,整个过程持续70 min,用脱脂奶粉(5%)阻断膜2 h。一抗CDKN1A稀释(1 ∶1 000)、GAPDH稀释(1 ∶1 000)在4 ℃下孵育过夜。HRP二抗稀释(1 ∶10 000)在37 ℃下孵育2 h、使用增强化学发光成像。利用BandScan进行灰度值分析。
1.2.4实时荧光定量聚合酶链反应(qRT-PCR)检测 使用1 ml TRIzol提取细胞总RNA。用 miScript SYBR Green PCR Kit试剂盒检测mRNA和miRNA的表达水平。大约3 μg的总RNA用于互补DNA(cDNA)的合成。使用2 μl的cDNA进行PCR扩增。反应条件为:95 ℃ 30 s ; 95 ℃ 5 s , 60 ℃ 32 s,40个循环; 95 ℃ 15 s, 60 ℃ 1 min, 95 ℃ 15 s,60 ℃ 15 s,运用2-ΔΔCt相对定量法进行数据分析。CDKN1A引物(F:5′-CCCGTGAGCGATGGAACTT-3′;R:5′CGAGGCACAAGGGTACAAGAC-3′);以GADPH作为对照 (F:5′-TCAAGAAGGTGGTGAAGCAGG-3′;R:5′-TCAAAGGTGGAGGAGTGGGT-3′);kshv-mir-k12-1-5p引物(F:5′-TGCGCATTACAGG AAACTGGGTG-3′; R:5′-CCAGTGCAGGGTCCGAG GTATT-3′;茎-环结构引物(stem-loop):5′-GTCG TATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATA CGACGCTTACAC-3′)。以U6作为对照(F:5′-CGCTTCGGCAGCACATATAC-3′;R:5′-AAATATGGAACGCTTCACGA-3′)。
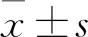
1.2.5细胞周期(PI法)测定 将kshv-mir-k12-1-5p mimics、inhibitor等(50 nmol/L)分别转染到SLK细胞中。48 h后获得2×105个细胞。用200 μl的细胞液离心5 min,用PBS洗涤细胞2次。随后,细胞在-20 ℃下用75%乙醇固定1 h,在4 ℃下预冷。用冷PBS再次清洗细胞。细胞周期分析试剂盒的使用方法参照生产厂家提供的说明书,将Rnase A溶液(20 μl)加入细胞,37 ℃下置于水浴中30 min,然后将PI染料溶液(400 μl)加入混合物中,4 ℃下在黑暗中孵育。采用流式细胞术检测,检测波长488 nm。(PI值=S+G2/G1+S+G2,其中PI为增殖活性指数。PI值越大,细胞周期进程越快,细胞周期时间越短,细胞增殖越快。)
2 结果
2.1 kshv-mir-k12-1-5p的转染水平将kshv-mir-k12-1-5p mimics、 inhibitor等分别转染到SLK细胞中,qRT-PCR结果显示,mimics组kshv-mir-k12-1-5p转染水平最高(4.43±0.07), inhibitor组kshv-mir-k12-1-5p转染水平最低(0.72±0.02)。各组转染水平的差异有统计学意义(F=2 971.42,P<0.01),见图1。

图1 qRT-PCR检测kshv-mir-k12-1-5p的转染水平
1:normal cell group; 2:mimics 组; 3:inhibitor组;4:mimics NC组; 5: inhibitor NC组; 与 mimics NC组比较:**P<0.01;与inhibitor NC组比较:##P<0.01
2.2 CDKN1A 3′UTR与kshv-mir-K12-1-5p的结合位点预测通过Mirbase(http://www.mirbase.org/)靶基因预测软件分析,从数百个靶基因中选择CDKN1A作为kshv-mir-K12-1-5p的靶基因验证,因为CDKN1A与细胞周期调控密切相关,是功能强大的细胞周期负性调控因子。图2显示了CDKN1A 3′UTR与kshv-mir-K12-1-5p的种子区序列(Seed sequence)的结合位点。
2.3 kshv-mir-k12-1-5p特异靶向CDKN1A293T细胞分别转染kshv-mir-k12-1-5p mimics+CDKN1A-WT、mimics NC+CDKN1A-WT、kshv-mir-k12-1-5p mimics+CDKN1A-MUT和mimics NC+ CDKN1A-MUT。24 h后测定荧光素酶的相对活性。kshv-mir-k12-1-5p mimics和CDKN1A-WT共转染后,相对荧光素酶活性降低(4.56±0.18);kshv-mir-k12-1-5p mimics和CDKN1A-MUT共转染后,相对荧光素酶活性无明显变化(7.11±0.17)。 mimics NC和CDKN1A-WT/ CDKN1A-MUT转染后相对荧光素酶活性无明显影响[(6.29±0.19)vs(7.04±0.21)]。各组荧光素酶的相对活性差异有统计学意义(F=28.31,P<0.01)。因此,CDKN1A是kshv-mir-k12-1-5p直接作用的靶基因。见图3。
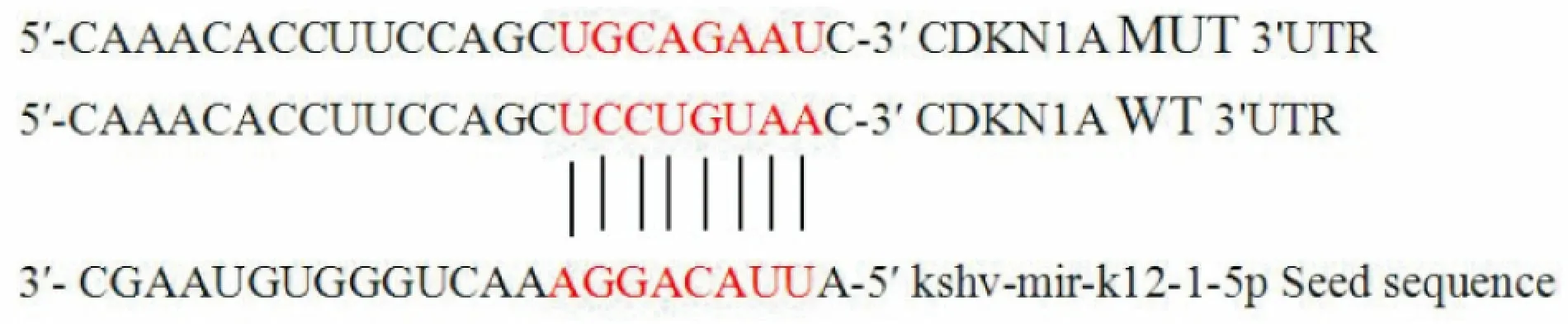
图2 CDKN1A 3′UTR与kshv-mir-k12-1-5p的结合位点

图3 各组相对荧光素酶活性
1:mimics+CDKN1A-WT组;2:mimics NC+CDKN1A-WT组;3:mimics+CDKN1A-MUT组;4:mimics NC+CDKN1A-MUT组;与mimics+CDKN1A-WT组比较:**P<0.01
2.4 kshv-mir-k12-1-5p负性调控靶基因CDKN1A的表达转染kshv-mir-k12-1-5p mimics后,SLK细胞中CDKN1A的蛋白表达水平显著低于其余4组;转染kshv-mir-k12-1-5p inhibitor后,SLK细胞中CDKN1A的蛋白表达水平显著高于其余4组;转染kshv-mir-k12-1-5p mimics后,SLK细胞中CDKN1A mRNA的表达水平显著低于其余4组;转染kshv-mir-k12-1-5p inhibitor后,SLK细胞中CDKN1A mRNA的表达水平显著高于其余4组。各组CDKN1A的蛋白水平(F=76.10,P<0.01)和mRNA表达水平(F=83.31,P<0.01)差异有统计学意义。因此,kshv-mir-k12-1-5p可以负调控靶基因CDKN1A的表达。见图4。
2.5 kshv-mir-k12-1-5P对SLK细胞周期的影响转染SLK细胞48 h后,用流式细胞术检测细胞周期分布情况。结果显示:与其余4组比较,kshv-mir-k12-1-5p mimics组PI值最高,G1期细胞数量最少,S+G2期细胞数量最多;与其余4组比较,kshv-mir-k12-1-5p inhibitor组PI值最低、G1期细胞数量最多、S+G2期细胞数量最少。各组PI值差异有统计学意义(F=220.01,P<0.01)。因此,kshv-mir-k12-1-5P可加速细胞周期进程,缩短细胞周期时间,促进细胞增殖,促进G1/S期转换。见图5。

图4 kshv-mir-k12-1-5p负性调控靶基因CDKN1A的表达
A:Western blot法检测结果图;B:CDKN1A蛋白相对表达量;C:CDKN1A mRNA 相对表达量;1:normal cell group; 2:mimics组; 3:inhibitor组;4:mimics NC组; 5: inhibitor NC组; 与mimics NC组比较:**P<0.01;与inhibitor NC组比较:##P<0.01
3 讨论
卡波西肉瘤是间叶源性高度播散的血管肉瘤[1]。其在中国新疆高发,主要见于维吾尔族和哈萨克族。KS与获得性免疫缺陷综合征(AIDS)、KSHV、血管生成细胞分化和遗传易感性有关。KSHV是导致人类恶性肿瘤的七种病毒之一,可引起人类宿主终身感染,是KS的重要致病因子。KSHV主要通过编码的miRNAs发挥作用,KSHV miRNAs对宿主细胞的周期及自身周期均有重要的调控作用,最终可以诱导肿瘤的发生[10]。
Catrina Ene et al[12]通过基因芯片分析了17例KS样本与3例正常皮肤样本中miRNAs的表达差异,结果显示:kshv-mir-k12-4-3p、kshv-mir-k12-1在KS中表达明显上调,Wu et al[9]通过KS及瘤旁组织中miRNAs表达差异谱分析得出:69个miRNAs在KS中表达明显上调,101个miRNAs表达明显下调。其中kshv-mir-k12-1-5p表达上调最为明显。因此高表达的kshv-mir-k12-1-5p可能与KS的细胞调控密切相关,但需要进一步验证。miRNAs 的功能研究关键在于其靶基因的确定。因为miRNAs通过抑制靶基因的功能来发挥生物学作用[11]。如KSHV感染的淋巴瘤细胞中KSHV miRNA调控的潜在靶基因在该病的发病机制中起关键作用[13]。
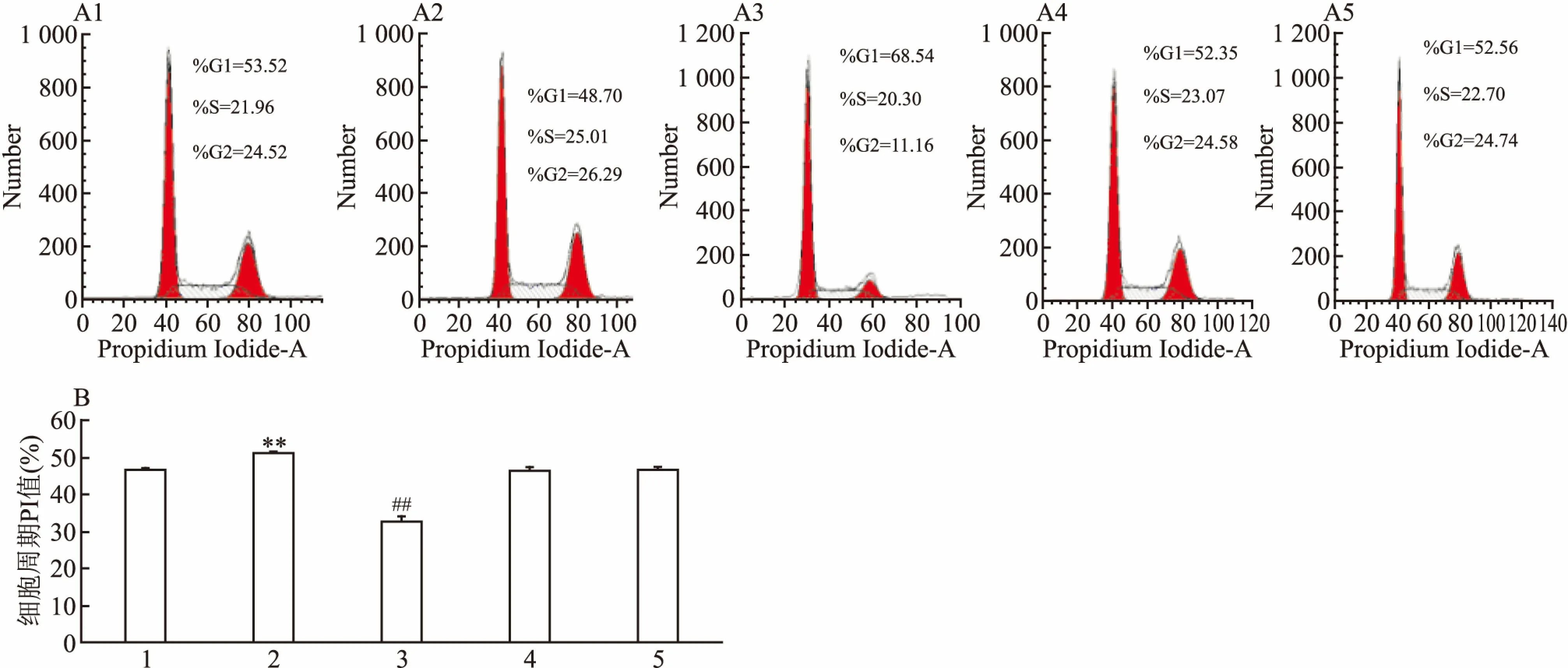
图5 kshv-mir-k12-1-5P对SLK细胞周期的影响
A:细胞周期分布;B:细胞周期PI值(%);1:normal cell group; 2:mimics组; 3:inhibitor组;4:mimics NC组; 5: inhibitor NC组; 与 mimics NC组比较:**P<0.01;与inhibitor NC组比较:##P<0.01
CDKN1A是细胞周期相关基因,位于6号染色体上。作为一种抑癌基因,在肿瘤中广泛低表达。细胞周期有两个重要的检查点:G1/S期和G2/M期。从细胞周期进程来看,G1/S期的检查点调控比G2/M期更为重要。CDKN1A主要作用于G1/S期检查点[14],抑制G1期向S期进展。与P53间接作用细胞周期不同,CDKN1A直接与 Cyclin / CDK复合蛋白或CDK蛋白激酶结合,负性调控细胞周期。当受到化学或物理刺激时,CDKN1A可以修复损伤的DNA,如果CDKN1A受到抑制,细胞没有完全修复直接进入细胞周期,复制错误遗传信息,将导致肿瘤的发生[15]。
本研究通过检测荧光素酶表达活性确定了kshv-mir-k12-1-5p可特异性地作用于CDKN1A-3′UTR的结合位点,CDKN1A是kshv-mir-k12-1-5p作用的靶基因。通过体外细胞实验发现: kshv-mir-k12-1-5p表达上调时,SLK细胞中CDKN1A 的蛋白和mRNA表达水平均明显下降;kshv-mir-k12-1-5p表达下调时,CDKN1A 的蛋白及mRNA表达水平明显升高,可见kshv-mir-k12-1-5p能负性调控 CDKN1A的表达。此外,本研究应用细胞周期PI法分析kshv-mir-k12-1-5p对KS细胞周期的影响,结果表明:kshv-mir-k12-1-5p过表达后KS细胞的G1期细胞数量明显减少,S期和G2期细胞数量明显增加;抑制kshv-mir-k12-1-5p表达后,KS细胞发生G1期细胞阻滞,S期和G2期细胞数量明显减少。因此kshv-mir-k12-1-5p可以促进KS细胞G1/S期转换,加速细胞周期的进程。下一步本课题组将进行体内动物实验,明确 kshv-mir-k12-1-5p和靶基因共同参与KS发生、发展的机制,为KS的治疗提供理论依据和技术支持。
综上,kshv-mir-k12-1-5p可能通过下调或沉默靶基因CDKN1A的表达发挥细胞周期调控作用。它具有促癌基因的功能,正性调控KS的细胞周期,缩短细胞周期时间,促进G1/S期转换。导致细胞周期调控紊乱,诱导KS的发生。而抑制kshv-mir-k12-1-5p的表达有可能成为KS的一个新的治疗靶点。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21 02:14:50
中日友好医院学报(2021年1期)2021-04-14 01:58:32
传染病信息(2021年6期)2021-02-12 01:52:20
山东医药(2020年9期)2020-05-20 01:12:16
国际呼吸杂志(2019年4期)2019-03-12 01:07:30
小哥白尼·野生动物画报(2017年8期)2018-01-16 17:58:51
小哥白尼·野生动物画报(2017年10期)2018-01-15 18:55:46
中华老年多器官疾病杂志(2016年7期)2016-04-28 08:43:05
癌症进展(2016年10期)2016-03-20 13:15:43
医学研究杂志(2015年11期)2015-06-10 06:44:03
