
外源性精胺抑制高尔基体应激减轻高糖诱导的心肌损伤*
2020-01-01 07:37范玉琪王跃虹邵毅英邵小婷徐长庆
中国应用生理学杂志 2019年5期
范玉琪, 王跃虹, 邵毅英, 邵小婷, 扈 敬, 徐长庆, 魏 璨
(哈尔滨医科大学病理生理学教研室, 哈尔滨 150086)
糖尿病是世界范围内一种常见代谢性疾病,其特征是由于胰岛素分泌障碍或胰岛素抵抗而引起的血糖浓度升高[1]。糖尿病性心肌病(diabetic cardiomyopathy, DCM)是指糖尿病患者发生的不能用冠心病、高血压性心脏病及其他心脏病解释的心肌病变[2]。在大多数情况下,DCM患者早期无明显病理改变和临床症状,随后发生心肌肥厚和纤维化等,进而引起心肌收缩和舒张功能障碍,最终可导致充血性心力衰竭(congestive heart failure, CHF)[3]。然而其确切的发病机制尚未阐明。Khullar等人的研究表明,过度的氧化应激被认为在DCM的发展中发挥关键作用[4]。
高尔基体(Golgi apparatus, GA)是重要的细胞器,其主要功能为将内质网合成的蛋白质进行加工、分类与包装。在氧化应激过程中,高尔基体通过调节自身相关基因(如糖基化酶或膜转运因子)的转录,影响膜 Ca2+/Mn2+ATPase的活性,改变膜表面活性成分等对自身结构及功能做出适应性调整,此即为高尔基体应激(Golgi apparatus stress, GAS)[5]。Yang Z等人的研究表明,高尔基体是一种β1-肾上腺素能信号的连接体,可以决定高尔基体微环境中的局部Ca2+浓度,从而影响原代心肌细胞的蛋白质合成和运输[6]。Malik S 等发现了一种新的G蛋白偶联受体(G protein-coupled receptor, GPCR)依赖性调节心肌肥厚的途径,它依赖于高尔基体磷脂酰肌醇4-磷酸(phosphatidylinositol 4-phosphat, PI4P)在核膜上的特异性磷脂酶Cε(PLCε)水解[7]。Luisa Jungk 等[8]在慢性心房颤动患者中也发现了高尔基体应激的存在。但是,在糖尿病心肌病(DCM)的发生过程中是否有高尔基体应激的参与,目前尚未见报道。
多胺是广泛存在于自然界中的多价阳离子烷基胺,包括精胺,精脒和腐胺。其中精胺在细胞的增殖、分化与凋亡中发挥重要作用[9]。本课题组的前期研究发现,多胺代谢紊乱参与大鼠心肌缺血/再灌注损伤和心肌肥大的发生,给予外源性精胺可发挥心肌保护作用[10]。因此,我们推测在DCM的发生过程中有GAS的参与,外源性精胺可能通过抑制GAS在糖尿病心肌病中发挥保护作用。本文就此问题进行研究,旨在为DCM的防治提供新靶点和新思路。
1 材料与方法
1.1 实验动物
60只雄性Wistar大鼠(200~250 g,购自哈尔滨医科大学实验动物中心),随机分成三组(n=20):正常对照组(Control),糖尿病组(T1D),精胺组(T1D+Sp)。本实验采用60 mg/kg STZ(溶于0.1 mol/L 柠檬酸-柠檬酸钠缓冲液)一次性腹腔注射,复制1型糖尿病模型;对照组注射相同体积的柠檬酸-柠檬酸钠缓冲液;精胺组连续两周每天注射精胺(5 mg/kg/d),然后进行STZ注射,之后每隔1 d注射精胺至12周。实验大鼠给予STZ前禁食12 h;注射完STZ后立即喂24 h 10%蔗糖水。注射 STZ一周后,大鼠尾静脉采血测定空腹血糖浓度(fasting blood glucose concentration, FBG),如果FBG≥16.7 mmol/L 则确定为糖尿病大鼠造模成功。12周后,整晚禁食,腹腔注射戊巴比妥钠30 mg/kg 麻醉,处死各组大鼠。血样收集到含有抗凝剂的试管中。分离心脏,一部分用2.5% 戊二醛固定,用于透射电镜观察。一部分储于-80℃,用于其他实验。
1.2 超声检测大鼠心肌功能
注射STZ后0周和12周,在3%戊巴比妥钠(30 mg/kg)麻醉下,将Wistar大鼠剃毛,用GEVIVID 7,10S探头,10 MHz进行彩色多普勒超声心动图二维成像与TDI(Tissue Doppler imaging, TDI)技术测定大鼠心脏结构功能参数:左心室收缩期内径(left ventricular diameter in systole, LVIDs)和左心室舒张期内径(left diastolic dimension, LVIDd)和射血分数(ejection fraction, EF)。所有设置均经过优化,以获得最大信噪比和二维图像。
1.3 测定心肌酶cTnT、CK-MB浓度的测定
将收集到的血样以4℃、3 000 r/min离心15 min,留取上清。利用试剂盒(南京建城生物工程研究所)测定血清中心肌肌钙蛋白(cardiac trophonin T, cTnT)、肌酸激酶同工酶(creatine kinase isoenzyme-MB, CK-MB),所有试验均按照试剂盒说明书进行。
1.4 细胞模型建立及其分组
H9C2系大鼠心肌细胞购自上海斯信生物科技有限公司。将该细胞在含10 % 胎牛血清(FBS)、100 U/ml 青霉素和100 mg/ml链霉素的 DMEM 培养基中培养。当细胞达到 70%~80% 汇合时(通常传至6~10代)进行实验。将H9C2细胞随机分成3组(n=8):正常对照组(Control),用10%的FBS-DMEM 培养;高糖组(HG),培养基中加入40 mmol/L葡萄糖;精胺组(HG+Sp)在10%的FBS-DMEM中加5 μmol/L的精胺预处理30 min后,换成含5 μmol/L的精胺和40 mmol/L高糖的10%的FBS-DMEM中继续共同培养48 h。接种时,大皿中细胞密度为1×108个/皿,12孔板的细胞密度为1×106cells/well。
1.5 Western blot检测蛋白的表达情况
取冷冻保存的心肌组织,液氮研磨,放入1.5 ml EP管中,加入含PMSF的RIPA裂解液裂解在冰上裂解20 min,4℃、13 500 r/min离心30 min,取上清制备组织蛋白提取液;处理48 h后各组心肌细胞,按类似方法处理同样留取上清,用 BCA 法测定蛋白含量。用12%的SDS-PAGE 电泳分离蛋白,根据目的蛋白分子量选择配制不同浓度的分离胶,上样,电泳,至溴酚兰抵达凝胶底部时,停止电泳。采用湿转法将蛋白转移至PVDF膜上,室温,5% 脱脂牛奶封闭 1 h 后,裁剪目的蛋白所在的条带。将目的条带分别置于抗体盒中,加入相应的一抗(GOLPH3、GM130、Cleaved Caspase3 按1∶1 000比例配制),4℃过夜。次日用TBST洗膜,然后加入稀释过的二抗(1∶10 000),室温孵育 1 h。TBST洗膜,ECL法进行显色。应用 Image J 图像处理软件对条带进行分析,测量灰度值,计算各组目的蛋白的相对表达量。
1.6 免疫荧光观察高尔基体结构的变化
将接种在载玻片中的心肌细胞洗涤,固定并透膜。然后,将细胞用特异性一抗(GOLPH3,比例为1∶150)在4℃下免疫标记过夜。用PBS洗涤后,将细胞与相应荧光二抗:山羊抗兔IgG(Rockland Immunochemicals Inc, Limerick, PA)和DAPI核复染剂孵育,然后利用荧光显微镜(Olympus IX 51,Center Valley, PA)进行观察。
1.7 统计学处理
2 结果
2.1 外源性精胺对TID大鼠心肌损伤的干预作用
与正常组相比,糖尿病组大鼠血糖明显升高,体重显著下降,证实糖尿病造模成功;CK-MB、cTnT明显升高;心肌超微结构损伤明显(肌丝断裂,润盘消失等);射血分数(EF)降低,左心室收缩期内径(LVIDs)和左心室舒张期内径(LVIDd)显著增加,说明糖尿病模型制备成功,大鼠出现糖尿病心肌病的改变外源性精胺处理可以减轻上述变化(表1,表2,图1)
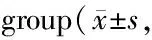
Tab. 1 Changes in blood glucose, body weight, ejection fraction and left ventricular function related parameters in each n=8)
TID: Diabetic group; TID+Sp: Spermine group; EF: Ejection fraction; LVIDs: Left ventricular internal diameter end-systolic; LVIDd: Left ventricular internal diastolic diameter
*P<0.05vscontrol group;#P<0.05vsT1D group
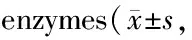
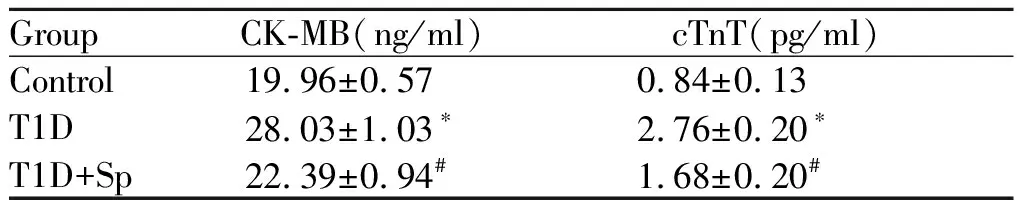
GroupCK-MB(ng/ml)cTnT(pg/ml)Control19.96±0.570.84±0.13T1D28.03±1.03∗2.76±0.20∗T1D+Sp22.39±0.94#1.68±0.20#
CK-MB: Creatine kinase-MB; cTnT: Troponin T
*P<0.05vscontrol group;#P<0.05vsT1D group
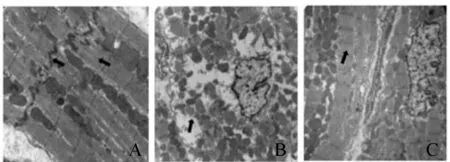
Fig.1Transmission electron microscopic observation of myocardial ultrastructure in each group (uncolored ×400)
A: Control; B: TID; C: TID+Sp
2.2 外源性精胺对TID大鼠心肌高尔基体应激的干预作用
分别以cleaved Caspase3,GOLPH3和GM130与β-tubulin的比值为参数,与Control组相比:T1D组cleaved Caspase 3和GOLPH3蛋白表达量显著增加(P<0.05),而GM130蛋白表达量显著下降(P< 0.05);与T1D组相比:T1D+Sp组cleaved Caspase3和GOLPH3蛋白表达量显著下降(P<0.05),而GM130蛋白表达量显著增加(P<0.05,图2,表3)

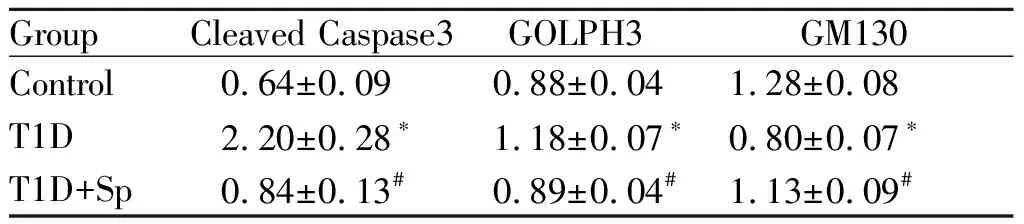
GroupCleaved Caspase3GOLPH3GM130Control0.64±0.090.88±0.041.28±0.08T1D2.20±0.28∗1.18±0.07∗0.80±0.07∗T1D+Sp0.84±0.13#0.89±0.04#1.13±0.09#
*P<0.05,**P<0.01vscontrol group;#P<0.05,##P<0.01vsTID group
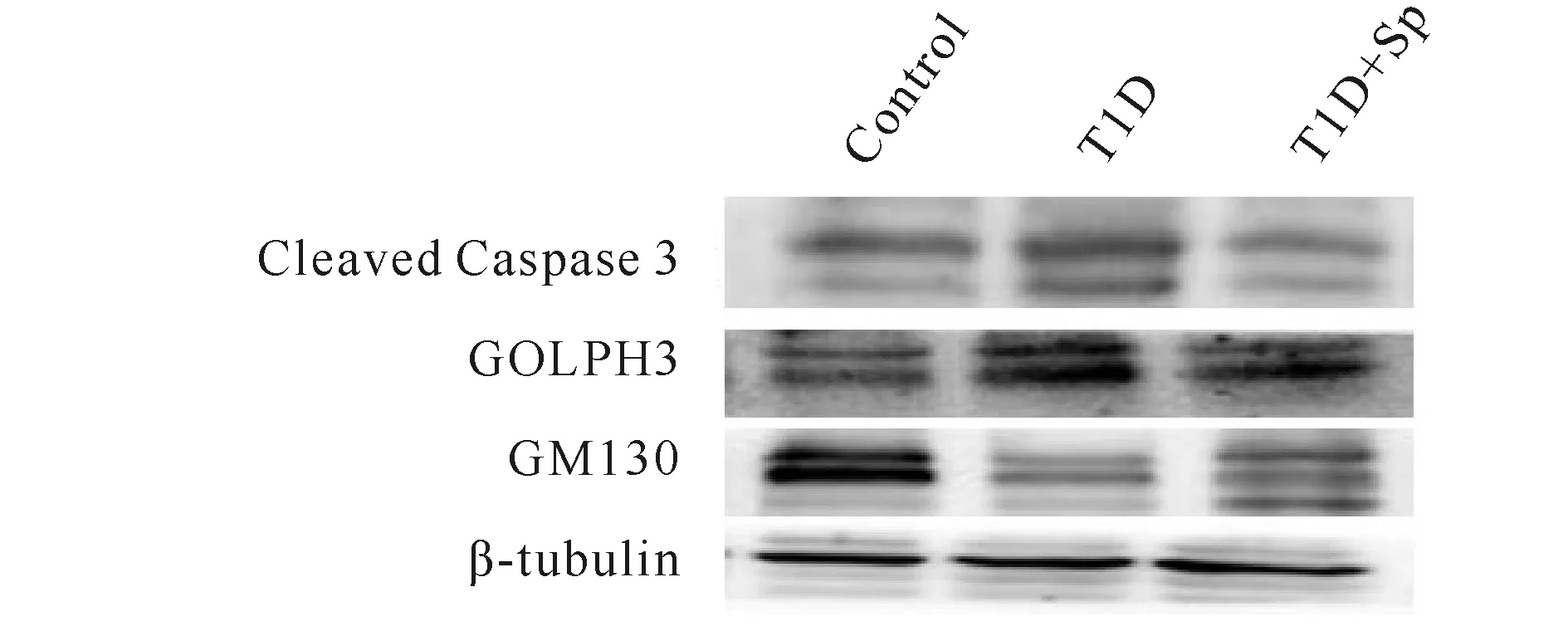
Fig.2Comparison of the expressions of golgi bodies and apoptosis-related proteins in myocardium of rats in each group (n=5)
2.3 外源性精胺对HG诱导的H9C2细胞高尔基体应激的干预作用
分别以cleaved Caspase3,GOLPH3和GM130与β-actin的比值为参数,与Control组相比:HG组cleaved Caspase3和GOLPH3蛋白表达量显著增加(P<0.05),而GM130蛋白表达量显著下降(P< 0.05);与HG组相比:HG+Sp组cleaved Caspase3,GOLPH 3蛋白表达量显著下降(P<0.05),而GM130蛋白表达量显著增加(P<0.05,图3,表4)
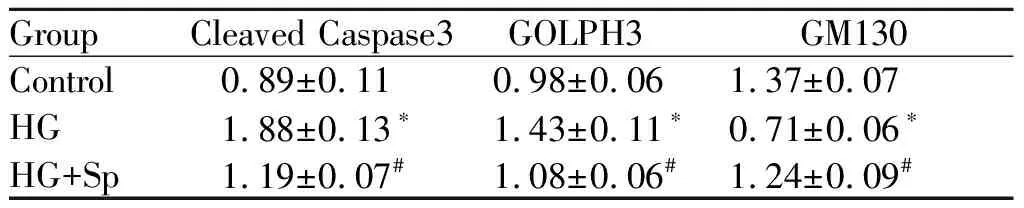
GroupCleaved Caspase3GOLPH3GM130Control0.89±0.110.98±0.061.37±0.07HG1.88±0.13∗1.43±0.11∗0.71±0.06∗HG+Sp1.19±0.07#1.08±0.06#1.24±0.09#
HG: High glucose group; HG+Sp: Spermine group
*P<0.05,**P<0.01vscontrol group;#P<0.05,##P<0.01vsHG group
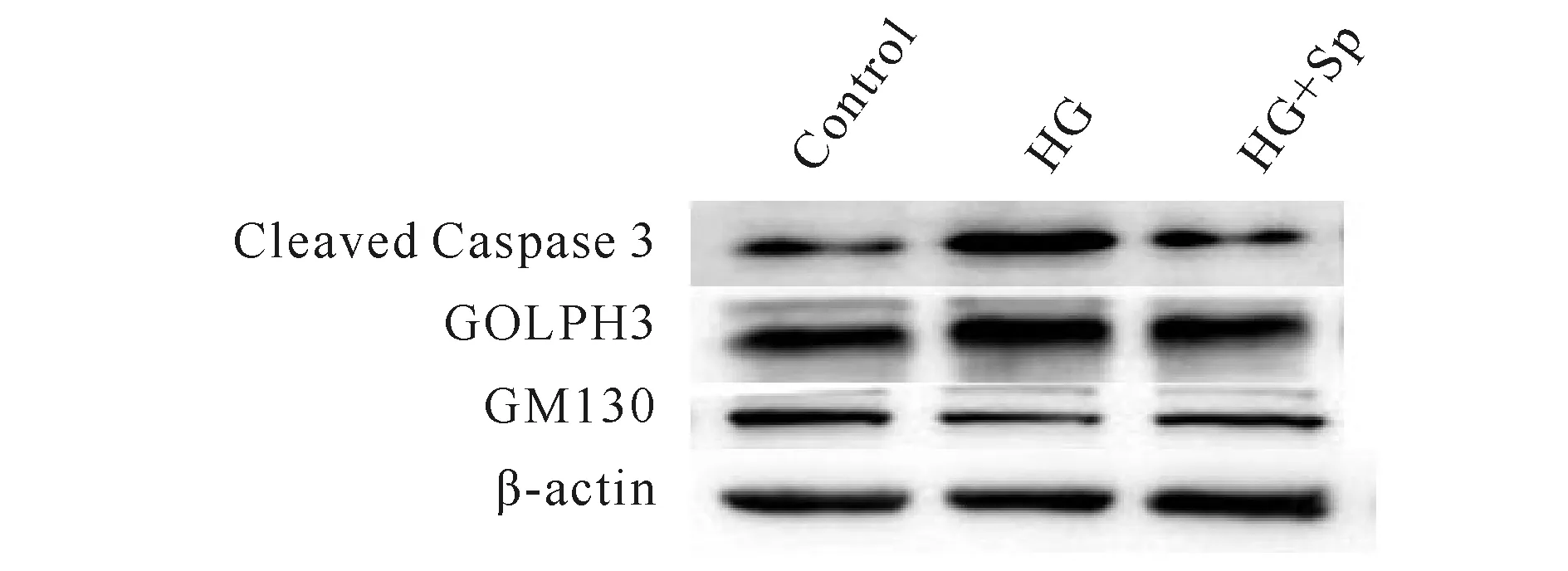
Fig.3Comparison of expressions of Golgi apparatus and apoptosis-related proteins in H9C2 myocardial cells in each group (n=5)
2.4 外源性精胺对HG诱导的H9C2细胞高尔基体碎片化的干预作用
对细胞质中的 GOLPH3 和细胞核 DAPI 进行双染。结果表明,在正常组的H9C2 细胞中,GOLPH3 标记的高尔基体为聚集在一起的浓密的带状结构;与正常组相比,HG处理组,高尔基体的由紧凑的囊泡样结构向胞浆弥散;加精胺处理后,高尔基体结构弥散程度减弱(图4)。
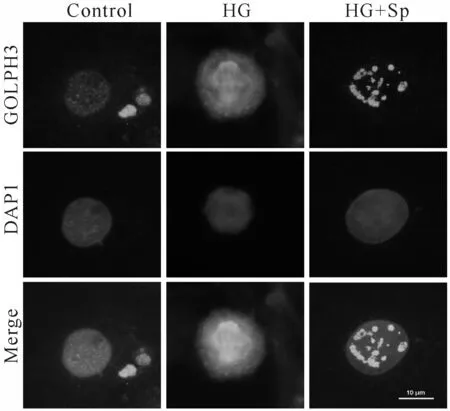
Fig.4Changes of Golgi apparatus in H9C2 myocardial cells of each group (Immunofluorescence ×400)
3 讨论
本文采用大剂量STZ注射成功复制了1型糖尿病(T1D)大鼠模型(血糖升高和体重减轻),模型组大鼠出现心肌酶释放增加,心肌收缩和舒张功能障碍及心肌超微结构损伤。这些结果提示DCM大鼠模型成功建立,为后期研究奠定了基础。
DCM的发生机制尚未完全阐明,一般认为与氧化应激、钙稳态失衡、线粒体障碍、炎症和纤维化等有关,其中过度氧化应激尤其关键[4]。氧化应激是一个多细胞器协同参与的复杂生理和病理过程,高尔基体作为关键细胞器同样参与其中。高尔基体除了对来自于内质网的内容物进行加工(例如,蛋白质水解、磷酸化、糖基化、磺化及脂类分类等)外,还参与离子稳态(特别是Ca2+)和细胞凋亡等的调控,这些均与氧化应激关系密切[11]。
GOLPH3是高尔基体来源的应激相关蛋白,位于高尔基体外膜,其表达水平与细胞内氧化应激水平呈正相关[12]。同时它还是一种磷脂酰肌醇-4-磷酸(PI4P)的效应物,在维持高尔基体结构和顺行转运方面起重要作用[13]。GM130(也被称为GOLGA2)是一种顺式高尔基体基质蛋白, 有助于维持高尔基体的带状形态结构特征。GM130磷酸化后将不再与高尔基体P115结合,是高尔基复合体分裂的原因之一[14]。强烈高尔基体应激可导致高尔基体的碎裂,进而引起高尔基体相关细胞凋亡[15]。
由于GM130和GOLPH3在高尔基体正常结构功能及高尔基体应激发挥重要作用,因此本实验检测了在DCM模型中这两种蛋白的表达。
在 DCM 大鼠心肌组织和高糖损伤的 H9C2 心肌细胞中,我们发现 GOLPH3蛋白表达显著升高,说明损伤后高尔基体应激的发生。而 GM130 的表达明显降低,GOLPH3 免疫荧光染色显示高尔基体结构由正常的紧凑囊泡状逐渐向胞浆弥散,都表明应激条件下高尔基体结构发生破坏。众所周知,Caspase3是细胞凋亡的共同执行者,其增加必然引起细胞凋亡增加[16]。上述实验结果,初步证实糖尿病心肌病(DCM)的发生有高尔基体应激的参加,并会促进凋亡的发生。
为了防治糖尿病心肌病(DCM),人们开展一系列研究。本课题组前期研究发现,DCM大鼠和高糖处理的心肌细胞均发生多胺代谢紊乱,内源性精胺减少。给予外源性精胺可延缓和减轻糖尿病心肌病的进展和损伤程度,其心肌保护机制主要通过抑制p38和JNK通路,减少活性氧的产生[17];增加CaSR表达,维持细胞内钙稳态[18];抑制内质网应激和线粒体凋亡通路[19]。
本文在上述研究成果的基础上,进一步观察外源性精胺能否通过抑制高尔基体应激而发挥心肌保护作用。为此,在实验分组时除了正常对照组和模型组外,还增加了(模型+精胺)组。本文结果显示,外源性精胺处理可以明显减轻DCM大鼠心脏和高糖处理的心肌细胞发生的各种病理变化。这些结果清楚地说明,外源性精胺可通过抑制高尔基体应激并减少细胞凋亡进而发挥对糖尿病心肌病的心肌保护作用。
需要强调的是,我们的实验结果只是初步证明高尔基体应激参与了糖尿病心肌病(DCM)的发生发展,外源性精胺的心肌保护作用与减轻高尔基体应激(GAS)有关。详细具体的机制还有待今后进一步研究。
猜你喜欢
昆明医科大学学报(2022年4期)2022-05-23
上海交通大学学报(医学版)(2022年3期)2022-05-05
中国典型病例大全(2022年9期)2022-04-19
中华养生保健(2020年4期)2020-11-16
保健文汇(2020年7期)2020-08-21
世界科学技术-中医药现代化(2020年2期)2020-07-25
世界科学技术-中医药现代化(2020年2期)2020-07-25
保健与生活(2020年13期)2020-07-24
上海党史与党建(2018年1期)2018-08-28
法制与社会(2017年22期)2017-09-04
