
携带miRNAs的MSCs来源外泌体在软骨退变损伤修复中的研究进展
2019-12-26 08:56:34静综述李章华审校
医学研究生学报 2019年12期
胡 静综述,李章华审校
0 引 言
随着人口老龄化和肥胖人群激增,关节软骨退变损伤,如关节炎(Osteoarthritis,OA)等已成为常见的慢性疾病之一,预计到2020年OA将成为全球第4位的致残原因[1]。OA表现为关节软骨降解和结构破坏,常年疼肿、僵硬使患者苦不堪言,严重影响生活质量[2]。间充质干细胞(mesenchymal stem cells,MSCs)是一类具有自我复制能力和多向分化潜能的多能干细胞,能向软骨分化治疗软骨相关疾病已成为共识[3]。其通过旁分泌和自分泌(内分泌)系统分泌多种细胞因子和生长因子,抑制局部免疫系统,参与受损组织的修复与再生[4]。外泌体是由细胞分泌的直径为30~150nm的囊泡,内含来源于细胞的相关蛋白质与核苷酸等生物分子,作为通讯的媒介对细胞间的交流有非常重要的作用[5]。MSCs来源的外泌体在免疫调节作用下可有效转运mRNA、miRNAs和蛋白质等生物活性物质,具备减少细胞凋亡、减轻炎症反应、促进基质合成和提高组织修复潜能等生物学功能[6]。众多研究表明,携带miRNAs的MSCs来源外泌体在软骨修复和再生中具有恢复细胞生物学活性、增加细胞数量和提高细胞免疫调节能力的作用[7-8]。本文就携带miRNAs的MSCs来源外泌体在软骨损伤退变性疾病修复与再生中发挥的作用作一综述。
1 软骨退变损伤治疗现状
软骨退变损伤十分常见,据流行病学显示,65岁以上中老年人群中50%以上有骨性关节炎,且随着年龄增长发病率会越来越高[9]。骨性关节炎通常是体育、娱乐活动以及衰老致关节软骨损伤引起的后遗症,其特征为关节软骨降解、半月板和韧带退变、软骨下骨增厚和骨赘形成,是造成中老年人骨关节疼痛和致残的主要原因[10]。软骨退变损伤目前尚无有效药物治疗,只能针对疼痛、肿胀和僵硬等症状进行对症处理,常见治疗方法有补充氨基葡萄糖、口服非甾体抗炎药、局部针灸、理疗、中药熏洗以及电刺激等[11],但这些治疗方法对软骨退变损伤的治疗效果未能尽如人意。近年来外泌体、miRNAs的研究取得了突破性的进展,为软骨退变损伤的修复提供了一种新思路。
2 外泌体
2.1外泌体来源最近的研究发现,胞外囊泡根据粒径大小可分为外泌体、微泡和自噬小体三种细胞外载体,功能各异,外泌体大多参与生理性的调控,微囊泡则更多介导病理性调节,至此外泌体发展到一个新的高度[12]。相对于其他组织,骨和软骨组织中外泌体的研究时间较短。1975年,首次在骨髓中发现细胞外膜颗粒,这表明多发性骨髓瘤细胞外囊泡与骨组织损伤之间可能存在联系[13]。1979年,通过显微镜检测到牙槽骨衍生的细胞外基质囊泡,这是首次发现正常骨衍生的胞外囊泡[14]。1980年,通过超微结构技术研究了成骨细胞衍生的基质囊泡。直到21世纪初,骨髓基质衍生的外泌体成为研究的焦点,其在向软骨细胞、成骨细胞等分化过程中起着重要的作用[15]。2017年以来,高通量、基因芯片等技术成熟,深入寻找外泌体内含的miRNAs及其相关功能成为研究的热点和前沿。
2.2外泌体的组成和功能外泌体的功能和生物学特性由其内在的含量决定。在外泌体成分中,有脂质、蛋白质和核酸三种主要成分,决定外泌体的特异性并将其与其他细胞外囊泡相区分[16]。外泌体的结构和分选很大程度上依赖于脂质,过去的几十年中,已对36个不同类别癌细胞的520多种脂质进行鉴定[17]。脂质通常富含外泌体膜,质膜中的主要非极性脂质是甾醇,是来自晚期体内高度富集的多泡体[18]。鞘磷脂是鞘质的主要成分,在外泌体膜的构建中发挥重要作用,还参与了货物的分选[19]。此外,鞘磷脂还富集于膜脂筏中,神经酰胺和二酰基甘油的下调介导外泌体处于平衡状态[20]。在miRNA分选中,中性鞘磷脂酶2是第一个被认为与该机制相关的分子;通过蛋白质组学分析,在所有哺乳动物外泌体中发现了多种蛋白质,如细胞骨架成分(微管蛋白,肌动蛋白,cofilin,profilin)、膜联蛋白(膜联蛋白Ⅰ,Ⅱ,Ⅳ,Ⅴ和Ⅶ)以及GTP酶家族成员rab7和rab11等。见图1。在外泌体蛋白质中,细胞质外泌体富集蛋白如Alix和TSG101,四跨膜蛋白如CD9、CD63和CD81是区分外泌体与其他细胞外颗粒的标志物,这些蛋白质的检测使研究人员能够快速评估外泌体的特征[21]。
外泌体还广泛存在于核酸中,编码和非编码RNA,单链和双链DNA均在外泌体中被发现。有研究显示,在哺乳动物细胞来源的外泌体中检测到1600个mRNA和700个miRNA,其中的miRNAs通常与细胞增殖、蛋白质合成和RNA转录后修饰密切相关[22]。此外,富含miRNAs的外泌体还能激活免疫系统,刺激T淋巴细胞、B淋巴细胞和树突细胞的释放,并参与T淋巴细胞和抗原递送细胞之间的相互作用[23]。在肿瘤生长、转移和耐药性的研究中发现,外泌体miRNAs在某些疾病中会发生特异性变异,可用于部分疾病的早期检测诊断[24]。上述可知,外泌体miRNAs对机体的调节是多系统的,其发挥作用的机制尚未完全可知,是目前研究的热点和难点。
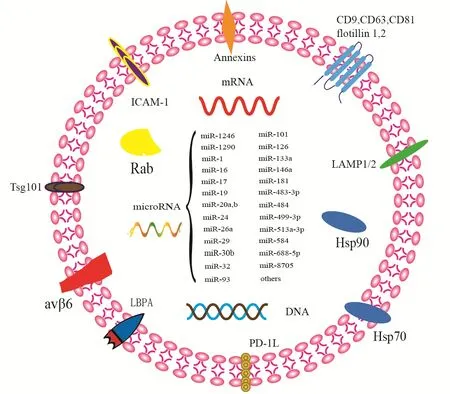
图1 外泌体的组成与功能示意图Figure 1 Composition and function diagram of exosomes
3 MSCs来源外泌体
MSCs是非造血基质干细胞,具有多向分化的潜能,能够分化为成骨细胞、肌腱细胞、软骨细胞、脂肪细胞等[25]。许多研究表明,MSCs通过自分泌和旁分泌释放多种细胞因子、趋化因子和生长因子介导组织修复[26]。Murphy等[27]在研究MSCs对软骨细胞的营养作用时发现,人骨髓MSCs通过分泌营养因子促进软骨细胞增殖和细胞外基质合成。随着蛋白质分离和蛋白质组学技术发展,发现MSCs分泌物的种类越来越多,包括细胞因子、生长因子、外泌体和miRNAs等[28],这些活性成份能促进细胞增殖、分化和生存,促进软骨生成和减少炎症反应[29]。MSCs来源外泌体内含MSCs的相关脂质、蛋白质和核苷酸等生物分子,在功能上延续了MSCs的特性。Zhang等[30]研究发现,在免疫活性大鼠软骨缺损模型中,人胚胎MSCs源性外泌体加速了新组织的填充,增强Ⅱ型胶原基质合成和硫酸化糖胺聚糖,外泌体处理的大鼠软骨和软骨下骨完全恢复,具有良好表面规则性的透明软骨、与相邻软骨完全粘合、细胞外基质沉积。相反,等渗盐水治疗的对照组中仅存在纤维性修复。此外,外泌体miRNAs的表达水平与关节炎高度契合,强大的miRNAs家族通过IL-1β、IGF、NF-κb等信号通路参与到炎症的反应中[31]。前述可知,MSCs源性外泌体对骨关节炎有调节作用,具体机制可能和内含的miRNAs和相应的通路密切相关。
4 软骨损伤相关的miRNAs及其信号通路
4.1 miRNAs家族与软骨损伤miRNAs是一类广泛存在的生物基因组,参与了生命中的一系列重要过程,如早期胚胎发育,细胞增殖、凋亡、死亡和脂肪代谢等[32]。miRNAs序列、结构和表达方式的特殊性,使其可能作为mRNAs编码蛋白质的调节因子,对基因表达、细胞周期调控等产生重要影响[33]。有研究发现,miR-140在斑马鱼、小鼠等的关节软骨中显示出特异性高表达,并且在骨关节炎关节软骨中的表达较正常关节软骨中的表达明显降低,这可能和一系列的信号通路和靶基因调控有关[34]。吴松寒等[35]用木瓜蛋白酶注射膝关节腔造成SD大鼠骨性关节炎模型,同时体外用IL-1β刺激骨性关节炎的软骨细胞模拟体外炎性环境,检测发现,miR-25对抑制软骨细胞凋亡过表达明显而对促进软骨细胞凋亡表达不足。Kim等[36]发现,骨关节炎患者中,miR-223的表达在OA早期比正常对照组显著升高,在OA软骨细胞中,PEX-16显著降低的同时miR-223表达增加;当沉默PEX-16的表达,细胞凋亡明显升高,同时过表达的miR-223促进人关节软骨细胞死亡并加重大鼠关节软骨的破坏。综上所述,miRNAs家族众多成员与软骨退变损伤密切相关,随着研究的深入其机制也越来越明了。
4.2 miRNAs在软骨损伤中的信号通路miRNAs家族可特异性表达于软骨组织中,并在OA发病进程中起着重要作用,通过影响多条信号传导通路及多种信号分子对OA的发病过程进行调控[37]。Mao等[38]研究发现,外源性miRNA-95-5p过表达在软骨的形成过程中可有效抑制组蛋白去乙酰化酶2/8,上调软骨表型特异基因二型胶原、sox9等。进一步研究发现,外泌体miRNA-95-5p可经HDAC调节组蛋白的修饰进而促进软骨形成,具体机制可能是抑制Ⅰ类HDAC来发挥作用,这在外泌体miRNA和HDACi在软骨形成的调节上是一个新的认识。Tao等[39]发现,滑膜MSCs过表达miRNA-140后来源的外泌体能够携带Wnt5a和Wnt5b,Wnt5a和Wnt5b可替代Wnt信号转导途径激活yes相关蛋白(yes associated protein,YAP)蛋白并增强软骨细胞的增殖和迁移能力,同时减少胞外基质分泌带来的不良反应。体内实验中此方法也成功阻止了大鼠模型中的OA发展。OA的进展中,IL-1β扮演着十分重要的角色,可通过促进软骨细胞变性和凋亡影响OA。Ma等[40]发现,用白细胞介素-1β预处理软骨细胞后,miRNA-10a-5P的表达量增加。同时荧光素酶活性检测发现,加入siRNA处理miRNA-10a-5P的同源基因HOXA1可逆转miRNA-10a-5P抑制剂对软骨细胞凋亡的挽救作用,动物实验同样证实了这一现象。参与软骨退变损伤的通路还有很多,诸如NF-κB、BMP、SOX9、IGF、P53、ER等,这些信号通路在OA过程中参与调节软骨细胞新陈代谢、细胞增殖、分化、凋亡,和细胞外基质的合成与降解,以及炎症反应等过程[41],共同组建了一个庞大的网络系统参与到软骨损伤的调节中。
5 携带miRNAs的MSCs来源外泌体是极具优势的新一代软骨损伤治疗剂
综合大量文献报道来看,MSCs治疗疾病具有广泛的适应证,关节软骨退变损伤是其中之一。虽然MSCs还未被批准用于人体试验,但在动物实验中已广泛用于软骨损伤的治疗[42]。MSCs治疗存在时间、浓度和生物活性衰退等问题,一经干预产生不良反应时不能被立即清除[43]。但外泌体不同,MSCs来源外泌体耐受性较直接用MSCs好,且具有最小的免疫源性和毒性风险[43]。Wang等[44]研究发现,对比滑膜MSCs和胚胎MSCs来源外泌体作用于手术造成的小鼠内侧半月板不稳模型。结果显示,外泌体组ColⅡ上调明显,ADAMTS5和蛋白聚糖下调较单纯EMSC明显。体内还证实了外泌体在炎性微环境中通过影响胞外基质的合成与降解来治疗骨关节炎,同时SO染色结果也支持外泌体组优于单纯细胞组。MSCs来源外泌体富含大量miRNAs,能够调控多条软骨合成分解代谢重要信号转导途径,这些miRNAs在介导MSCs源性外泌体的功效中发挥重要作用,如miRNA-140、miRNA-92等可通过调节软骨增值和软骨形成来治疗软骨损伤[45]。Tao等[39]通过miRNA-140过表达MSCs,来源外泌体激活YAP蛋白促进软骨细胞的增殖和表行维持,且纯化的外泌体所需的量更小,免疫原性和癌变可能较单纯细胞小。促炎细胞因子等快速上调可导致基质降解和软骨损伤,MSCs来源外泌体可调节受损软骨区域的炎性环境,是重要的软骨再生治疗手段[46]。此外,MSCs外泌体的蛋白质组富含生物化学活性管理酶,可恢复体内平衡到最重要的活动细胞和组织微环境。MSCs来源外泌体通过细胞间交流释放生物活性因子促进胶原合成、抑制凋亡、刺激细胞增殖和传递免疫调节信号,招募或重组组织修复和再生所需要的细胞[47]。综上,MSCs来源外泌体含多种miRNAs,相互之间经多条信号转导发挥治疗作用,目前是软骨损伤最具前景的治疗方式。
6 结 语
多年来,细胞释放的双层磷脂膜囊泡不被认为具有生物学意义,随着细胞外囊泡(外泌体或EV)作为生物活性分子载体被识别,改变了人们的看法。这些细胞释放的载体可携带细胞相关的货物传递到局部或远处目标并执行特定的生物学功能,随着各类型的细胞外囊泡鉴定及生物学功能、货物装载、释放途径、靶向机制和囊泡加工复杂性升级,该领域的维度已经扩展[48]。MSCs来源外泌体在包括软骨组织在内的许多组织中均有修复和再生作用,近年来分子生物学技术发展解密了更多miRNAs的功效,携带miRNAs的MSCs来源外泌体在软骨损伤修复极具前景。但是用于临床还有一些问题亟待解决,如胞外囊泡尺寸的多样性怎么规范,不同尺寸具有不同的生物学功能,介导的细胞——细胞通信系统远超出现有认识;胞外囊泡是一起发挥作用还是各个间协同作用还不得而知,需要开发一种单囊泡分析工具进一步研究[49]。综上所述,外泌体在医学领域的应用极具前景,但还有一系列问题尚需深入研究。
猜你喜欢
临床肝胆病杂志(2023年9期)2024-01-06 10:44:19
精准医学杂志(2023年5期)2023-04-29 18:03:53
学苑创造·A版(2022年5期)2022-05-19 05:42:43
昆明医科大学学报(2022年2期)2022-03-29 00:52:18
色谱(2021年9期)2021-09-06 09:47:36
昆明医科大学学报(2021年5期)2021-07-22 07:32:50
老年医学研究(2021年6期)2021-03-09 06:34:18
科学导报(2019年40期)2019-09-03 04:36:04
汉字汉语研究(2019年2期)2019-08-27 00:47:56
国际呼吸杂志(2019年8期)2019-04-29 09:15:14
