
短期增水对高寒草甸补播草地植被群落和土壤养分的影响
2019-12-20 01:34李兴福丁成翔蒋福祯林长存苏德荣
草地学报 2019年6期
刘 禹, 李兴福, 丁成翔, 蒋福祯, 林长存*, 维 拉, 苏德荣
(1. 北京林业大学草业与草原学院, 北京 100083;2. 青海大学畜牧兽医科学院, 青海 西宁 810016; 3. 青海大学农林科学院,青海 西宁 810016)
全球气候剧烈变化及过度放牧造成三江源高寒草甸严重退化,这不仅危害农牧民生活,更导致当地水土流失、土壤裸露、物种丰富度减少等一系列生态问题[1]。为了控制草地退化,修复草地生态环境,青海三江源地区进行了退化草地补播。水分是限制草地植被生长的关键因子,对植物种子的萌发,幼苗生长,以及繁殖均有重要影响[2-3]。由于全球CO2浓度和平均气温持续升高,全球水分时空格局正在发生巨大改变,近50年来青藏高原地区冰川退缩加剧,气候变暖[4],降水呈逐渐增加趋势[5]。水热条件是影响植物群落的主要因素,降水变化导致土壤肥力、草地群落物种分布、物种多样性与初级生产力出现不同程度的改变[6]。降水变化不仅对干旱半干旱地区生态系统过程和功能影响显著,而且也可能影响相对湿润的地区[7]。王长庭等[8]在青藏高原高寒草甸的降水模拟试验表明,增加生长季降水有利于提高植被生物量、物种多样性和群落均匀度,以及影响禾本科植物的相对重要值。许庆民等[9]对海北高寒草甸的研究发现,增水当年植被物种丰富度和生物多样性指数得到显著提高,第2年物种数目和多样性指数均降低。武建双等[10]研究发现,随着生长季降水累积,藏北高寒草地地上生物量、物种丰富度、Shannon-Wiener指数和Pielou指数呈增加趋势。李长斌等[11]在高寒草地的研究结果表明,随着降雨强度增加,植被盖度增加,Pielou指数出现显著差异。李凤滋等[12]在高寒草甸进行多年增水的结果表明,土壤总碳、土壤总氮增加,表层土壤铵态氮总体有增加的趋势,硝态氮总体有减少的趋势。杨新宇等[13]的结果表明增水在降低土壤可提取有机碳含量的同时增加土壤微生物量碳。
然而,当前关于降水变化对植被群落和土壤养分的影响多集中在天然草原,对于人工草地的研究局限在草地生产力[14-15],由于青藏高原草地类型多样,环境气候差异较大[16]及水分增加量、增加时期的差异,使得草地生态系统对于水分状况改变的响应机制相当复杂,在同一地点进行多梯度增水的研究是很有必要的。鉴于此,本研究以青藏高原三江源区退化补播草地为研究对象,进行多梯度增水试验,分析降水变化处理下草地生态系统的植被群落和土壤养分变化,以期为全球气候变化背景下高寒草原对降水格局变化的响应机理提供理论参考。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原三江源自然保护区内,试验样地位于青海省河南蒙古自治县(北纬34°05′~34°56′,东经100°53′~102°16′),海拔3 800 m。当地全年有6~7个月气温低于0℃,多年平均气温-1.3℃~1.6℃,最暖月均气温9.2℃~11.5℃,最冷月均气温-9.9℃~-14.6℃,无霜期为5~33 d,全年结冻期161~186 d。生长季多年(1997—2018)平均降雨量为466.42 mm,年蒸发量1 414.6 mm。境内年日照时数2 551.8~2 577.2 h,平均日照百分率59%,太阳年辐射总量139.7~152.5 kcal·cm-2。5月日均气温稳定通过3℃时,牧草开始返青,9月后日均气温降至3℃时牧草基本停止生长,生长期仅为150~170 d。
研究区植被为高寒草甸,试验地为围封的补播草地,面积50 m × 50 m,补播前该地区土壤受鼠害危害严重(植被盖度30%~50%),优势植物为苔草(Carexspp.),细叶亚菊(Ajaniatenuifolia)、委陵菜(Potentillachinensis),风毛菊(Saussureajaponica)和矮生嵩草(Kobresiahumilis)。补播方式为机械化免耕补播,时间为2017年5月上中旬,补播后不再进行后续改良措施,主要补播草种为垂穗披碱草(Elymusnutans)和草地早熟禾(Poapratensis),补播量为15~22.5 kg·ha-1,补播草种比例为1∶1。土壤质地以中壤土为主,土壤容重0.93~1.14 g·cm-3,孔隙度56%~64%,pH值为7.5~8.1,透气蓄水性能良好。2018年5月灌溉前随机选取9个1 m x 1 m的样方进行样地基本情况调查,其中土壤有机质(Soil organic matter,SOM)含量为74.81 g·kg-1;土壤全氮(Total nitrogen,TN)含量为6.22 g·kg-1;土壤总磷(Total phosphorus,TP)含量为1.22 g·kg-1;有效磷(Available phosphorus,AP)为38.17 mg·kg-1;铵态氮(Ammonium nitrogen,NH4+)为41.67 mg·kg-1,硝态氮(Nitrate nitrogen,NO3-)含量为12.72 mg·kg-1。
1.2 试验设计
试验前下载并整理当地近21年(1997—2017年)植物生长季(5—9月)平均降水量,以此作为试验增水处理基准。试验设置5种增水处理,每个增水处理包括6个3 m×3 m的小区,各小区之间设置3 m的缓冲带和过道。增水量分别为对照(Control check,CK),增水0 mm;W1:增水量为29.15 mm,为5—9月多年平均降水量的6.25%;W2:增水量为58.30 mm,为5—9月多年平均降水量的12.5%;W3:增水量为116.61 mm,为5—9月多年平均降水量的25%;W4:增水量为233.21 mm,为5—9月多年平均降水量的50%。增水起始日为2018年5月15日,每种增水处理7 d进行1次增水[18],共增水18次,每次增水量等分各处理总增水量。若计划增水日有自然降水则试验降水顺延。
图1 试验年(2018年)样地生长季降水量和大气温度Fig.1 Precipitation and temperature in study site
灌溉用水来自样地邻近的河流,灌溉方式为微喷带喷灌,通过降低流量(25~34 L·h-1·m-1)增加单次灌溉时长来减少径流。小区放置2块太阳能板为设备提供电能,试验地气象数据和地表及地下温度使用天圻智能生态站和土壤墒情仪(型号WSS0G10A;IST.HRG C-110S,东方润泽生态公司)获取。通过Galcon GSI (New)灌溉控制器对样地灌溉进行实时控制。
1.3 样品采集和测定方法
6—8月每月月末不灌溉的日期采集样品,在每个小区内完全随机选取3处1 m×1 m的样方刈割植被地上部分,记录植物种类、高度、株丛密度、分盖度、总盖度和频度等,用于植物群落多样性计算[17]。在刈割后的样方采集土壤,使用环刀法采集0~20 cm的土样进行土壤含水量(Soil moisture content,SM),容重(Soil bulk density,BD)和孔隙度(Soil porosity,SP)的分析。用五点法采集0~20 cm的土壤样品进行土壤养分测定[19]。
采用烘干法测定土壤含水量[19];土壤pH值采用pH计测定。采用重铬酸钾外加热法测定土壤有机质(Soil organic matter,SOM)含量[19];采用凯氏定氮法测定土壤全氮(Total nitrogen,TN)含量[20];采用钼锑抗比色法测定土壤总磷(Total phosphorus,TP)含量[21];有效磷(Available phosphorus,AP)用Olsen法测定[19];铵态氮(Ammonium nitrogen,NH4+)用浸提—靛酚蓝比色法测量[22];硝态氮(Nitrate nitrogen,NO3-)用紫外分光光度法测量[23];亚硝态氮(Nitrite nitrogen,NO2-)用重氮偶合分光光度法测量[24];微生物碳(Microbial carbon,Bc)采用熏蒸法测量[25]。
采用物种丰富度指数Richness、Shannon-Wiener指数、Simpson指数和Pielou指数分析群落物种多样性;为了研究增水处理群落结构的影响,选用了2种补播植物(垂穗披碱草,草地早熟禾),2种优势原生植物(委陵菜,细叶亚菊)计算重要值指数(Important value,IV)。具体公式[26]如下:
丰富度指数:Richness=S
(1)
式中:S表示表示出现在某一草地类型中的物种数
Shannon-Wiener指数:H=-∑(PilnPi)
(2)
式中:Pi为此物种个体数占总个体数比例

(3)
式中:Pi为此物种个体数占总个体数比例
Pielou指数:Pielou=-∑(PilnPi)/lnS
(4)
式中:S为群落中的总物种数,Pi为此物种个体数占总个体数比例
重要值IV =(RA + RC + RF +RH)/4
(5)
式中:IV(Important value)为重要值,RA(Relative abundance)为相对多度,RC(Relative coverage)为相对盖度,RF(Relative frequency)为相对频度,RH(Relative height)为相对高度
1.4 数据分析
使用单因素分析测定同月不同增水处理下土壤含水量(SM)等土壤因子及植被地上生物量(Biomass)、盖度(Cover)、物种多样性指标等植被群落特征的差异;通过去趋势对应分析(Decentred correspondence analysis,DCA)计算得到排序轴梯度长度小于3,采用冗余分析法(Redundancy analysis,RDA)对土壤因子与植物数据矩阵进行排序分析。通过逐步回归分析(Stepwise regression)得到植被特征与土壤因子的关系。单因素和多因素方差分析采用SPSS 20.0统计分析软件,冗余分析用R3.5.3。绘图使用0riginPro 2015和R ggplot2包。土壤容重(Soil bulk density,BD)、孔隙度(Soil porosity,SP)和亚硝态氮数据仅用做冗余分析。
2 结果与分析
2.1 生长季不同增水处理下的植被特征
2.1.1地上植被生物量和盖度 6,7,8月均是W4处理植被地上生物量最高(图2A),其中7,8月增水处理与对照差异显著。与生物量类似,3个月的植被盖度同样是W4处理最高(图2B),除8月外,其余2个月4组增水处理的盖度均显著高于CK处理。
CP是消化道疾病中的常见病和多发病,分为肿瘤性和非肿瘤性息肉[5]。该病的病因尚不明确,有研究[1]显示,沿海地区渔民患病率较高,并且海上作业时间越长,CP患病率越高;海上作业<5年CP患病率为15.7%,5~10年为28.6%,>10年为37.2%。可能与其生活习惯和工作环境的特殊性有关,如长期海上作业饮食节律失常,新鲜蔬菜水果相对缺乏,饮酒吸烟等。
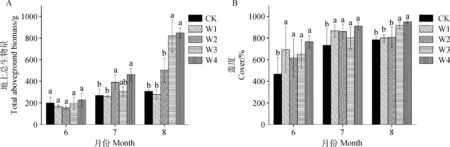
图2 各增水梯度生长季各月份(6—8月)植被地上生物量和盖度Fig.2 Aboveground biomass and cover of vegetation in each month (6—8) under different water addition treatments注:不同小写字母表示同一月份不同增水梯度差异显著(P<0.05),下同Note:Different lowercase letters indicate significant differences between the water addition treatments in the same month (P<0.05). The same as below
2.1.2植被物种多样性 植被群落多样性是植被群落特征的典型代表,可以很好地反映外部变化对群落中各物种生长发育和群落演替的影响。由图3可知,6月各处理间Shannon-Wiener指数、Simpson指数、Pielou指数均不存在显著差异,除W3外,其余增水处理丰富度指数均大于CK。7月增水处理与对照的4种指数均没有显著差异。8月各处理的Shannon-Wiener指数、Simpson指数和Pielou指数出现先减后增的趋势,在W4处理下达到最大值,但与CK处理没有显著差异(P>0.05);物种丰富度CK处理最大,并显著大于W2处理(P< 0.05)。
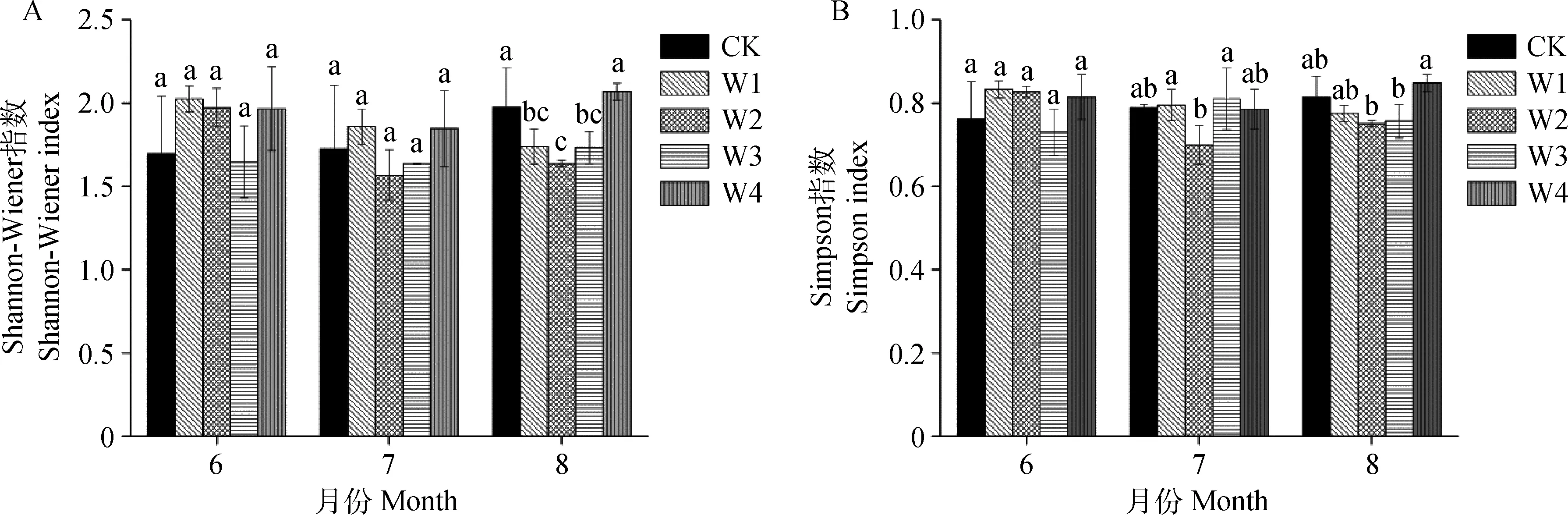

图3 各增水梯度各月份植被多样性Fig.3 Plant diversity in each month under different water addition treatments
2.1.3植被重要值 重要值是度量植物种群的关键指标,综合反应了植物物种在群落中的重要程度。由图4可知,生长季初期增水处理对垂穗披碱草的重要值影响较小,但显著降低了另一补播草种草地早熟禾的重要值,提高了细叶亚菊的重要值(图4A)。在生长季中期和末期,增水处理对垂穗披碱草的影响开始体现,增水处理下垂穗披碱草的重要值高于对照组,末期W4处理下的垂穗披碱草重要值显著高于其他处理;此时增水处理对草地早熟禾无显著影响(图4B,图4C)。

图4 各增水梯度各月份植被重要值Fig.4 Important values of vegetation in each month under different water addition treatments
2.2 草地生长季不同增水处理下土壤理化性质
2.2.1土壤含水量 对照处理土壤含水量为19.22%,4组增水处理平均土壤含水量为21.68%,含水量最高的为W4,为22.52%,最低是W1,为21.22%(图5)。从土壤含水量动态变化来看,6月份W4土壤含水量远大于其余4组处理,即增水首月土壤含水量对增水量的反馈较为明显;7月份CK处理土壤含水量远小于增水处理。8月份试验各处理差异较小,增水对土壤含水量的影响逐渐弱化。

图5 不同增水处理各月份土壤含水量Fig.5 Soil moisture in each month under different water addition treatments
2.2.2土壤pH 增水处理与对照处理间土壤pH差异不显著,随着增水量的增加,土壤pH呈现先降后增的趋势。6月份4个增水梯度与CK没有显著差异;7月份pH值呈现先减后增的趋势,CK土壤pH最高,W2处理土壤pH最低,两者差异显著;8月份各处理没有显著差异(表1)。
2.2.3土壤有机质、全氮、全磷 7月份W2处理下土壤SOM含量最高(图6A),为196.92 g·kg-1,显著高于同月其他4个处理(P< 0.05)。6,8两月各处理间SOM均不存在显著差异。整体上TN含量W2最高,但与CK处理无显著差异(图6B)。6,7,8三个月土壤TP含量变化呈单峰曲线,随增水量增加先增后降,都在W1处理下有最大值,且8月份W1的TP含量显著高于CK(图6C)。

表1 增水对6,7,8月土壤pH的影响Table 1 Effect of water addition on soil pH in different months
注:不同小写字母表示在同一月份不同增水处理中差异显著(P<0.05);ns表示P>0.05,差异不显著;*表示P<0.05,差异显著;**表示P<0.01,差异极显著;下表同
Note:Different lowercase letters indicate significant differences between the water addition treatments in the same month (P<0.05);ns indicates the difference is not significant.*indicates the difference is significant.**indicates the probability atP<0.01,and the difference is high significant. Similarly for the following tables
2.2.4土壤速效氮 从表2可以看出,增水处理下,6月份铵态氮和硝态氮含量在W2处理时含量最低,在W1和W3处理下较高,并显著高于其他处理。7月份是W3处理时铵态氮含量最高,8月份各处理间铵态氮和硝态氮不存在显著性差异(表2)。
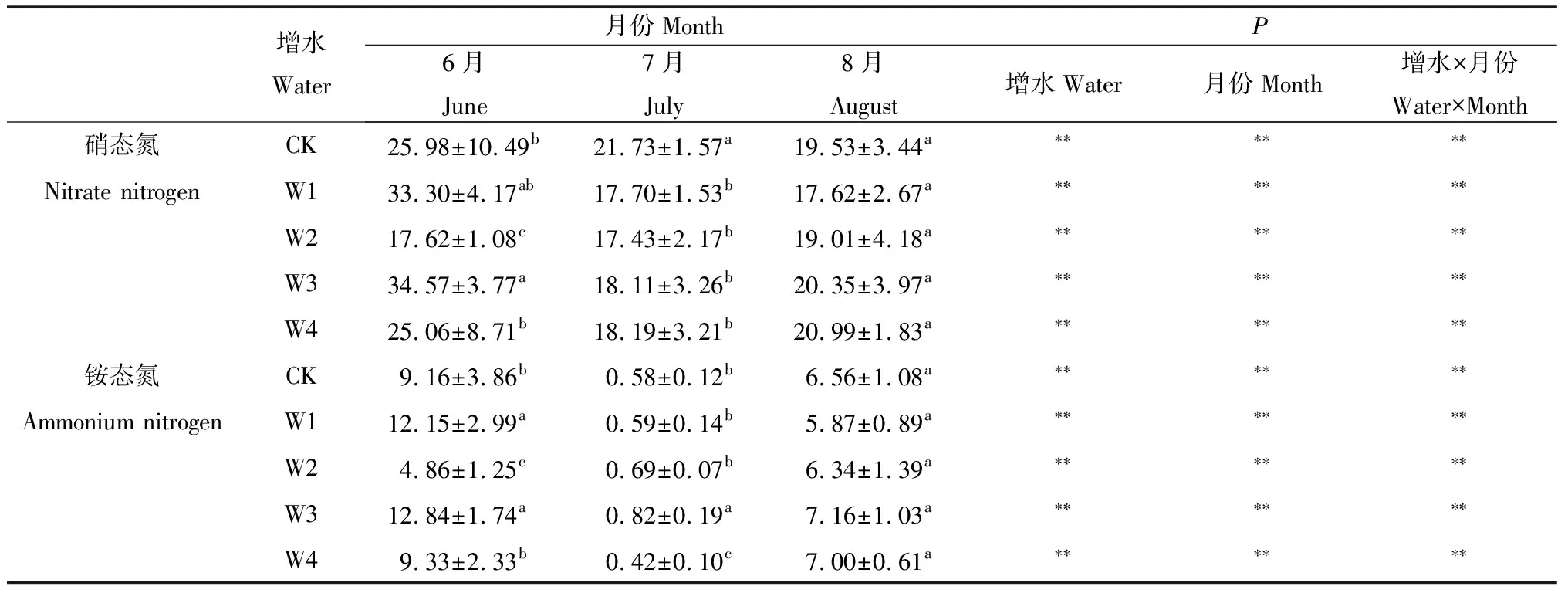
表2 增水对6,7,8月土壤速效氮的影响Table 2 Effect of water addition on soil available nitrogen in different months (mg·kg-1)

图6 各增水梯度不同月份SOM,TN,TP含量差异Fig.6 Differences in carbon,nitrogen and phosphorus contents in different months under different water addition treatments
2.2.5土壤微生物碳 6月份微生物碳呈先增后减的趋势,W2处理下土壤微生物碳含量最高,并且显著高于CK(P<0.05);7,8两月均是W4处理下微生物碳含量最高(P<0.05),且均显著高于CK(表3)。

表3 增水对6,7,8月土壤微生物碳的影响Table 3 Effect of water addition on soil microbial carbon in different months (mg·kg-1)

表4 增水对6,7,8月土壤有效磷的影响Table 4 Effect of water addition on soil available phosphorus in different months (mg·kg-1)
2.3 植被特征与土壤因子的关系
2.3.1植被特征与土壤因子的冗余分析排列 根据RDA的结果,前2个排序轴植被特征与土壤因子关系累计解释量达90.73%,由此可知第1,2轴能较好地反映植被群落特征与土壤因子的关系,且主要由第1轴决定。
植被群落特征均沿着第1轴排列,除了生物量外,其他均在轴的右侧;土壤养分因子中TN、硝态氮,有效磷,TP均沿第1轴排列,而土壤SOM、微生物碳沿第2轴排列,土壤含水量也沿第1轴排列,表明植被群落的多样性、生物量和生长主要受土壤氮、磷元素和土壤含水量不同程度的影响。由图7可知土壤含水量与植被生物量位于同一方向且夹角较小,表明土壤含水量与植被生物量呈显著正相关(P<0.05),与Shannon-Wiener指数、Simpson指数、Pielou指数、物种丰富度及植被盖度呈负相关。土壤养分因子中,土壤含水量与速效磷、土壤SOM、TN、TP、亚硝态氮等夹角小于90°,呈正相关;与微生物碳、铵态氮、硝态氮夹角大于90°,呈负相关。不同处理组的排布差异表明在高寒草甸不同增水量对土壤养分、土壤物理性质和植被群落特征的影响不同。
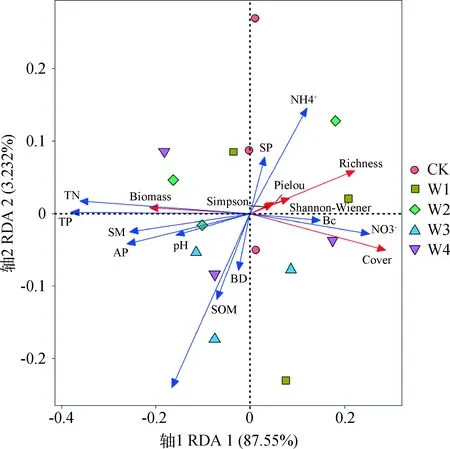
图7 植被特征与环境因子的冗余分析排序Fig.7 Redundancy analysis ordination of vegetation and environmental factors注:图中蓝色线段代表土壤因子,红色线段代表植被特征;Biomass为植被生物量、Cover为植被盖度、Richness为物种丰富度指数、Shannon-Wiener为Shannon-Wiener指数、Simpson为Simpson指数、Pielou为Pielou指数;SM为土壤含水量、BD为土壤容重、SP为土壤孔隙度、pH为土壤pH值、SOM为土壤有机质、TN为土壤全氮、TP为土壤全磷、Bc为土壤微生物碳、AP为土壤有效磷、NH4+为土壤铵态氮、NO3-为土壤硝态氮、NO2-为土壤亚硝态氮Note:The blue line represents soil factors;The red line represents vegetation characteristics. Biomass (Plant biomass),Cover (Plant cover),Richness (Richness index),Shannon-Wiener (Shannon-Wiener index),Simpson (Simpson index),Pielou (Pielou index),SM (Soil moisture content),BD (Soil bulk density),SP (Soil porosity),SOM (Soil organic matter),TN (Total Nitrogen),TP (Total phosphorus),Bc (Microbial carbon),AP (Available phosphorus),NH4+ (Ammonium nitrogen),NO3- (Nitrate nitrogen),NO2- (Nitrite nitrogen)
2.3.2植被特征与土壤因子的回归分析 通过线性回归分析(表5)发现,生长季6-8月植被生物量、盖度均与土壤因子成线性关系,且两方程中土壤含水量系数均显著。Shannon-Wiener指数与土壤因子的线性关系不显著,而Simpson指数、Pielou指数和物种丰富度均无法构成方程。生长季初期(6月)的多样性指数中仅有物种丰富度与土壤含水量的线性关系显著。
3 讨论
3.1 增水对高寒草甸植被特征的影响
植物群落特征受到植被群落类型、土壤养分含量等因素的影响,已有研究在探讨不同干扰因素对植被群落特征的影响时得出的结果不尽相同[27-28]。水分是影响植被群落类型、多样性及生长繁殖的关键因子。王志鹏等[29]研究表明植被生物量随着增水量增加而增加,本研究结果与其一致,即增水能够提高植被地上生物量。增水处理提高植被生物量的原因一方面是增水促进了土壤养分与水分耦合,最大限度地提高了植物所需养分;另一方面增水显著提高了垂穗披碱草和草地早熟禾的生物量,由图4可知这2类植物在群落中重要值较高,这2类植物发生变化对群落整体影响较大,从而提高了整体植被生物量。
本试验总体上增水处理的Shannon-Wiener、Simpson、Pielou等多样性指数均得到了提高,表明在青藏高原增水能够一定程度地增加高寒草甸物种多样性指数,这与李文娇等[30]和白春利等[31]在短花针茅荒漠草原上得到的结果有所不同,这可能是由于荒漠草原与高寒草原气候、土质、植被类型等空间异质性造成的。武建双等[10]研究发现藏北高原高寒草地Shannon-Wiener指数分布于0.92±0.05 到2.19±0.07之间,随生长季降水减少而降低,这与本研究生长季初期(6月)的结果一致。赵新风等[32]的结果虽然也表明植物生物多样性与年降水量显著正相关,但当水资源饱和时可能抑制非优势种生长,导致多样性降低,这也许是本试验增水处理后期(8月)物种丰富度指数呈下降趋势的原因。在增水处理后期,土壤含水量增加导致资源供应不对称,而试验地优势草种(垂穗披碱草,草地早熟禾)为禾本科,植株高大,生物量多,在土壤养分的争夺中占优势,群落中不同植物的地上、地下部分竞争力度改变,导致物种丰富度降低[33]。邓建明等[34]也发现高寒草甸群落中不同物种养分吸收能力和资源分配策略的差异较大,另外不同物种之间对群落下层光的竞争也是导致群落物种多样性指数降低的主要原因之一[35]。
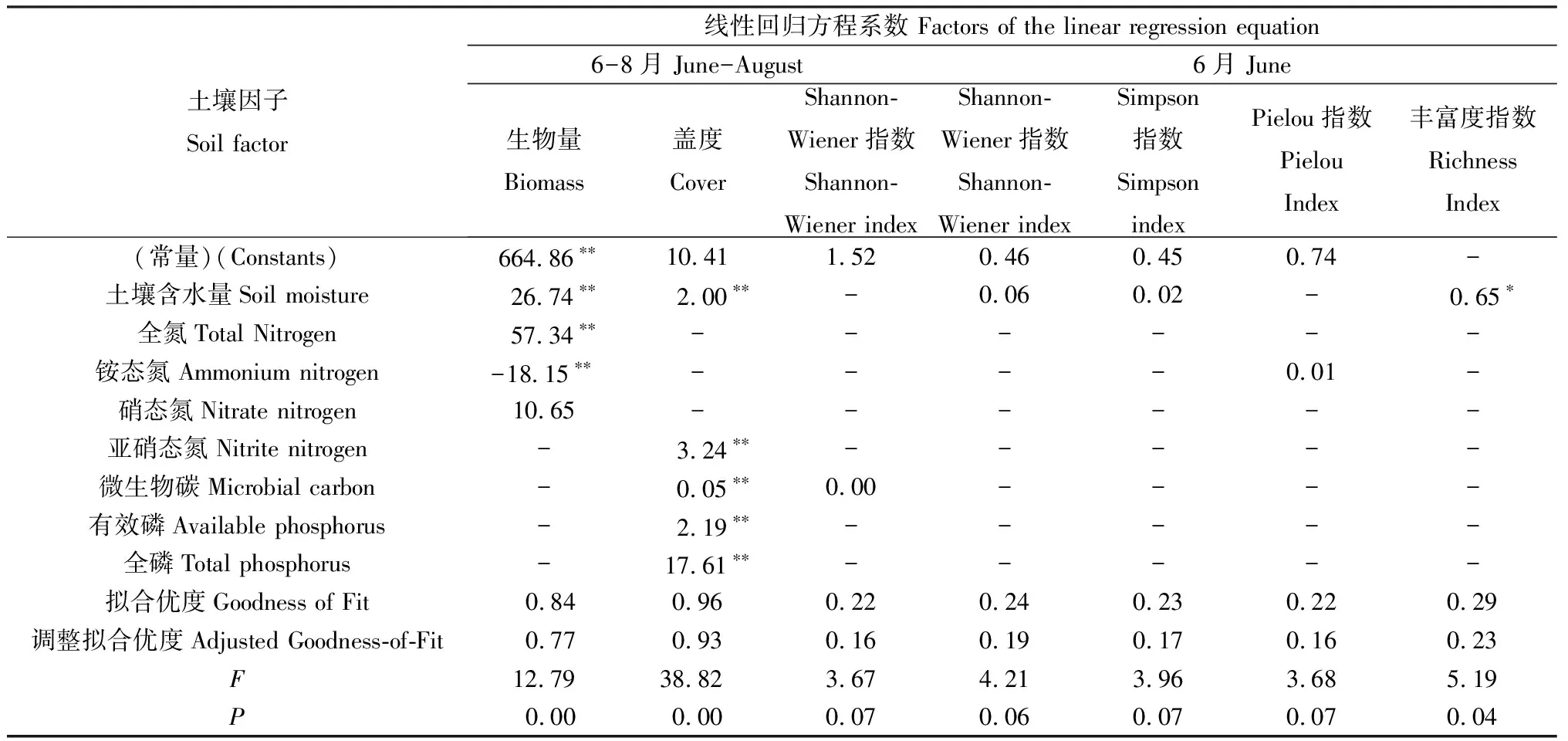
表5 植被特征与土壤因子的线性回归方程Table 5 The linear regression equation of vegetation characteristics and soil factors
注:*和**分别表示在0.05和0.01水平差异显著
Note:*and**indicate significant and high significant difference at the 0.05 and 0.01 level,respectively
3.2 增水对高寒草甸土壤因子的影响
有研究[36-37]在高寒草甸的试验中发现增水导致土壤温度降低,SOM分解速度减慢,表层土壤SOM累积,本试验也得出增水对土壤SOM累积有一定的促进作用,在增水12.5%(W2)处理下土壤SOM增加最多,增水量继续增加土壤SOM含量下降,说明生态系统中土壤总SOM含量短期内受增水量的变化影响有限。魏金明等[38]在内蒙古典型草原的研究发现,与不增水相比,增水处理土壤pH值从6.45提高到6.92,本试验结果与其相似,土壤pH与土壤含水量呈正相关。土壤水分增加有助于微生物所需养分基质的流通,提高微生物的分解能力。土壤微生物碳主要受土壤微生物数量影响,只占土壤全碳很小一部分,但却是土壤中最为敏感的因子,能够即时有效地对外部因素作出反馈。本试验中土壤含水量与土壤微生物碳呈正相关,土壤含水量增加土壤微生物含量增大。
已有研究[39]多认为增水后土壤含水量对硝化和矿化作用影响显著,且可以通过影响植物吸收氮的生物过程进而影响土壤氮循环。Liu等[40]在高寒高原冻土的研究结果表明氮和土壤含水量存在显著正相关;Luo等[41]对全球陆地的研究综述发现氮与年降雨量显著相关;Li等[42]在青藏高原高寒草甸的研究结果发现增水与对照处理的土壤全氮含量不存在显著差异,本试验结果与其一致,究其原因可能是增水显著增加了土壤含水量,有氧硝化在水饱和状态下受限,反硝化作用随水分增加而增强,即铵态氮随水分增加而增加,硝态氮减少[43]。土壤含水量增加导致土壤淋溶作用增强,尤其是硝态氮比铵态氮更容易随土壤淋溶丢失,本研究中7月份CK处理下硝态氮含量最高也证明了这一点。有研究[44]表明植被群落的分布和多样性与土壤因子有显著的相关关系。植被分布对土壤中氮、磷、有机质等营养物质的含量有显著影响,高寒草甸土壤因子对植被群落特征也具有重要的反馈作用,土壤全氮、铵态氮、硝态氮等养分与植被生物量有着紧密联系。
4 结论
增水处理显著提高了高寒草地植被地上生物量和盖度,且增水较多的处理组生物量较高。生长季初期增水显著提高了物种多样性指数,但是在生长季中期和后期多样性指数出现不同程度的下降。生长季前期增水提高了草地早熟禾的重要值,中、后期显著提高了垂穗披碱草的重要值。生长季初期和中期增水可以有效提高土壤含水量。结果表明人工增水对植被生长季初期缺水有明显缓解作用。土壤铵态氮、硝态氮、微生物碳、有效磷等速效养分对短期增水的反馈较为明显,增水可以有效提高土壤微生物碳、铵态氮含量。因此,在高寒草甸进行短期增水处理可以显著提高植被生物量和盖度,在50%的增水处理下影响最为明显,各梯度增水处理对不同土壤养分类型的影响不一,但总体上促进了土壤养分的累积。
猜你喜欢
今日农业(2020年19期)2020-12-14
种子(2019年8期)2019-09-11
时代农机(2018年11期)2018-03-17
环境污染与防治(2016年12期)2016-03-13
电源技术(2016年9期)2016-02-27
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
中国氯碱(2014年10期)2014-02-28
中国烟草学报(2012年3期)2012-04-10
植物营养与肥料学报(2011年4期)2011-10-26
