
短期增温对长江源区高寒沼泽草甸植物-土壤C,N化学计量及季节变化特征的影响
2019-12-20 01:27刘永万尹鹏松张景然
草地学报 2019年6期
刘永万, 白 炜, 尹鹏松, 冯 月, 张景然
(兰州交通大学环境与市政工程学院, 甘肃 兰州 730070)
近年来,由于工业的发展以及人类生活水平的快速提升,CO2及其他温室气体剧增,由此导致的全球气候变暖已引起世界各国的广泛关注[1]。据政府间气候变化委员会第五次评估报告显示,在过去130年来全球平均气温升高了0.85℃[2]。青藏高原作为世界上海拔最高的地区,因其严酷而脆弱的环境条件,对气候变化十分敏感[3]。王朋岭等[4]研究发现青藏高原地区平均气温以每十年0.4℃的速率增加,且青藏高原地区气温增加幅度明显高于同期北半球平均增温幅度。气温的持续升高导致青藏高原产生了一系列的生态问题,冻融周期的变化、气候暖干化趋势等直接影响着生态系统的物质循环,进而改变植物-土壤系统的化学计量特征。
碳(C)是植物有机质的主要成分,约占植物干重的1/2[5]。生态系统中植被的C主要来源于植物通过光合作用固定大气中的CO2;C含量反映了植物的光合碳同化能力,是植物C储量的一种度量。氮(N)是蛋白质、叶绿素、核酸等的一个重要组成部分,是植物生长所必需的重要营养元素之一。C和N作为植物生长所必需的养分元素,虽然其含量在特定植物种的特定生长阶段具有一定的变异性,但生物体具有保持其自身C,N元素组成相对稳定的能力[6],而这种相对稳定状态容易受到外界环境胁迫而被破坏[7],从而影响植物的生长、土壤C积累动态和N养分限制格局[8]。
到目前为止,基于增温对我国高海拔冻土区域高寒草地方面的研究多集中在植物群落组成、物种多样性和生物量等[3,8-9],对其化学计量学方面研究较少,且得出的结果各不相同。羊留冬[7]等人模拟气候变暖发现贡嘎山峨眉冷杉幼苗在增温处理后C平均含量和C/N均无显著变化,N含量显著降低;江肖杰[10]等人则发现增温并没有改变植物叶片的C,N含量,却使其土壤N含量、叶片C/N呈增加趋势;彭阿辉[6]等研究发现增温增加了植物地上部分C/N,但对不同植物C,N含量影响并不一致。上述研究多集中于植物领域,而对植物-土壤C,N营养元素传递与调节机理的研究还比较薄弱,对增温处理下,高寒草地植物、土壤化学计量比之间的关系及其规律更是鲜有报道。沼泽草甸是高寒草地生态系统的三大主要类型之一,下垫面为季节性冻土或永久冻土,温度变化会对土壤的物理性质、水分含量以及养分状况产生巨大影响,从而影响植物生长及其C,N含量。藏嵩草(Kobresiatibetica)是青藏高原风火山地区高寒沼泽草甸的优势种,其生长状态及C,N含量的改变都会对生态系统造成巨大的影响。因此,本研究以青藏高原腹地长江源风火山地区高寒沼泽草甸优势种藏嵩草为研究对象,设立增温样地,比较分析不同增温幅度下长江源区草地植物与土壤C,N生态化学计量比的变化与相互关系,为揭示植物-土壤C,N化学计量特征对温度变化的响应和调控机制提供科学依据。
1 材料与方法
1.1 试验地位置及植被概况
研究区域位于高寒沼泽草甸分布较为典型的青藏高原腹地风火山地区,海拔4 600~4 800 m,相对高差200~300 m,南北宽约40 km,整体呈北西西向延伸,属于可可西里山系的东南支[11]。属于高原大陆性气候区,寒冷干燥,冻结期 9月至翌年4月,年平均气温—5.3℃,年均降水量270 mm,且80%的降雨发生在植物的生长季(5-9月),年均蒸发量1 478 mm[12-13]。选取的样地位于34°43′43.9″ N,92°53′34.1″E,海拔4 754 m,其植物群落主要由湿中生、湿生多年草本植物群落构成。优势种为藏蒿草(Kobresiatibetica)、小蒿草(Kobresiahumilis)、针茅(Stipacapillata)、羊茅(Festucaovina)和粗喙苔草(Carexscabrirostris)等高寒植物。
1.2 试验设计
2015年7月,在研究区域内,随机选取植被相对分布均匀一致的区域,设置2种模拟增温处理:T1,增温 1.5~2.5℃;T2,增温 3~5℃。以自然样地为对照(CK),每种处理设置5个重复。
模拟增温的方式采用采用国际冻原计划(International Tundra Experiment,ITEX) 所采用的被动式增温法——开顶式气室法(Open-top chambers,OTC),建造开顶式小暖室,小室为正六边形圆台状结构,六面均为有机玻璃,透光率95%,小室的高度分别为40 cm和80 cm,并保证小室的上开口均为60 cm,圆台的斜边与地面的夹角均为60°。同时,采用EC-TM温湿度传感仪(Decagon Decices Inc.,Pullman,WA)记录空气温度与湿度。
1.3 研究方法
1.3.1取样方法 于2018年的6月(返青期)、8月(结实期)和9月(枯黄期),在每个处理样地内各随机选取10株藏嵩草,连同根一起取得完整的植株样品,将其周围枯枝和凋落物收集并分别装进信封袋,做好标记。同时采集植物根部周围层0~10 cm的土壤装入自封袋,做好标记。带回实验室的植物洗掉根部泥土后快速阴干,将地上、地下分开装袋,做好标记。于65℃烘箱内烘干至恒重,研磨、过筛用于有机碳和全氮的测定。同时,将土样阴干、研磨、过筛用于有机碳和全氮的测定。
1.3.2测定项目与方法 有机碳测定采用重铬酸钾外加热法;全氮测定采用半微量开氏法,经EHD20型Digi Block智能样品消解炉(莱伯泰科有限公司)消解后,由半微量蒸馏装置蒸馏,后经硫酸滴定测定[14]。
1.3.3数据分析 使用Origin2018进行数据制图,在SPSS 25中对各项数据进行整理和分析。采用单因素方差分析(One-Way ANOVA)对植物-土壤C,N含量及其化学计量比进行差异显著性检验,并采用Sparman相关系数法评价不同指标之间的相关性。
2 结果与分析
2.1 模拟增温的效果
由于温室的阻挡作用,增温棚室内风速降低,空气湍流减弱,使热量不易散失,加之玻璃纤维被太阳辐射中红外线穿透的能力较好[15],所以室内温度升高。在整个植物生长季内,沼泽草甸的平均气温分别比CK提高了3.1℃(T1)和5.5℃(T2),其中7月月平均气温达到最大值。根据以往研究,青藏高原地区平均气温以每十年0.4℃的速率增加[4],就这种趋势而言,本研究采用的增温系统在未来80~140 a,增温效果都是显著的(图1)。

图1 5-10月月平均气温Fig.1 Average temperature from May to October注:CK,自然样地;T1,增温 1.5~2.5℃;T2,增温 3~5℃,下同Note:CK,natural sample;T1,warming 1.5~2.5℃;T2,warming 3~5℃,the same as below
2.2 增温对植物地上部分C,N的影响
增温使得植物地上部分C含量减少,但只有结实期T2处理下差异显著(P<0.05),T1处理组相比CK减少了0.1%~3.2%,T2处理组相比CK减少了0.8%~9.8%,表明增温降低了植物地上部分C含量,但低增温对其影响并不显著,随着增温幅度的增加,会加剧这种效果。对增温处理后植物地上部分N含量来说,增温显著降低了返青期植物地上部分N含量(P<0.05),增加了结实期植物地上部分N含量,对9月枯黄期植物地上部分N含量并无明显影响(图2)。
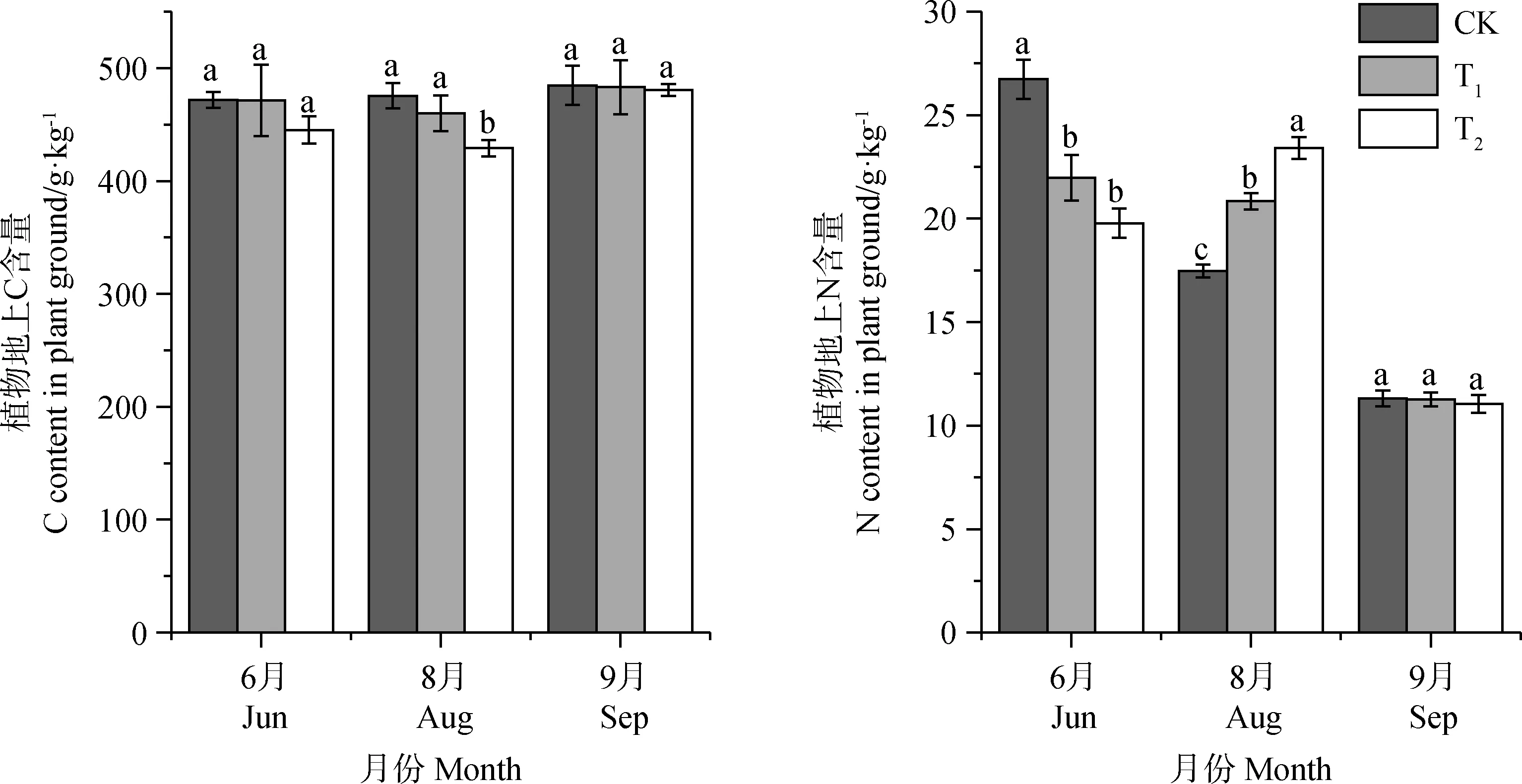
图2 不同增温幅度对植被地上部分C,N含量的影响Fig.2 The effect of different temperature increase range on the aboveground part of C and N content in different seasons注:不同小写字母表示不同增温幅度下的差异显著性(P<0.05),下同Note:Different lowercase letters indicate significant differences between different temperature increase range at the 0.05 level,the same as below
2.3 增温对植物枯落物C,N的影响
增温处理下6月返青期植物枯落物C含量显著增加,T1处理下增加了5.2%,T2处理下增加了15.8%。而8,9月枯落物C含量明显减少,T1处理组相比CK减少了8.2%~10.1%,T2处理组相比CK减少了1.7%~5.9%。对于增温处理后的枯落物N含量来说,增温显著降低了其N含量,T1处理组相比CK减少了9.2%~19.8%,T2处理组相比CK减少了6.1%~28.1%(图3)。
图3 不同增温幅度对植被枯落物C,N含量的影响Fig.3 The effects of different temperature increase ranges on the content of C and N in litter of different seasons
2.4 增温对植物根系C,N的影响
增温在6月返青期对植物根系C含量影响并不显著。T1和T2处理下8,9月根系C含量均显著低于对照(P<0.05),T1处理下其根系C含量相比CK降低了13.2%~25.1%;T2处理组相比CK降低了19.4%~25.7%,表明增温处理明显减少了植物根系C含量,但在返青期表现并不显著(图4)。
植物根系N含量对增温的响应有所不同,T1处理组显著降低了8月结实期植物根系N含量(P<0.05),使其减少了14.6%,对其他时期植物根系N含量影响并不显著。T2处理组在6月返青期、8月结实期显著增加了植物根系N含量(P<0.05),使其增加了10.1%~13.1%,在9月枯黄期则有一定程度的降低,但这种影响幅度较小,统计结果也表明差异并不显著。表明低增温减少了结实期植物根系N含量,对其他时期并无显著影响;高增温则促进了植物根系N含量。
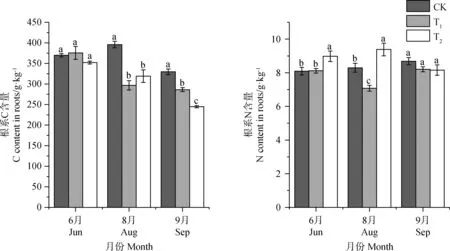
图4 不同增温幅度对植被根系C,N含量的影响Fig.4 The effect of different temperature increase range on the content of C and N in roots of different seasons.
2.5 增温对表层土壤C、N的影响
与CK相比,T1处理组明显促进了土壤表层有机碳含量,使其增加了12.2%~34.7%,T2处理同样促进了土壤有机碳含量,但返青期与CK之间差异并不显著,T2处理组相比CK使之增加了3.6%~214.0%。表明增温对土壤表层有机碳含量有明显的促进作用(图5)。
增温处理后,就土壤表层全氮含量来言,T1,T2处理下返青期土壤全氮含量均显著低于CK(P<0.05),分别减少了17.2%和18.7%;T1处理显著促进了8,9月土壤全氮含量(P<0.05),使其相比CK增加了17.2%~32.0%;T2处理同样表现为促进作用,但9月并不显著,使其增加了0.17%~99.2%。表明增温处理减少了返青期的土壤表层全氮含量,其他时期使土壤表层全氮含量明显增加。
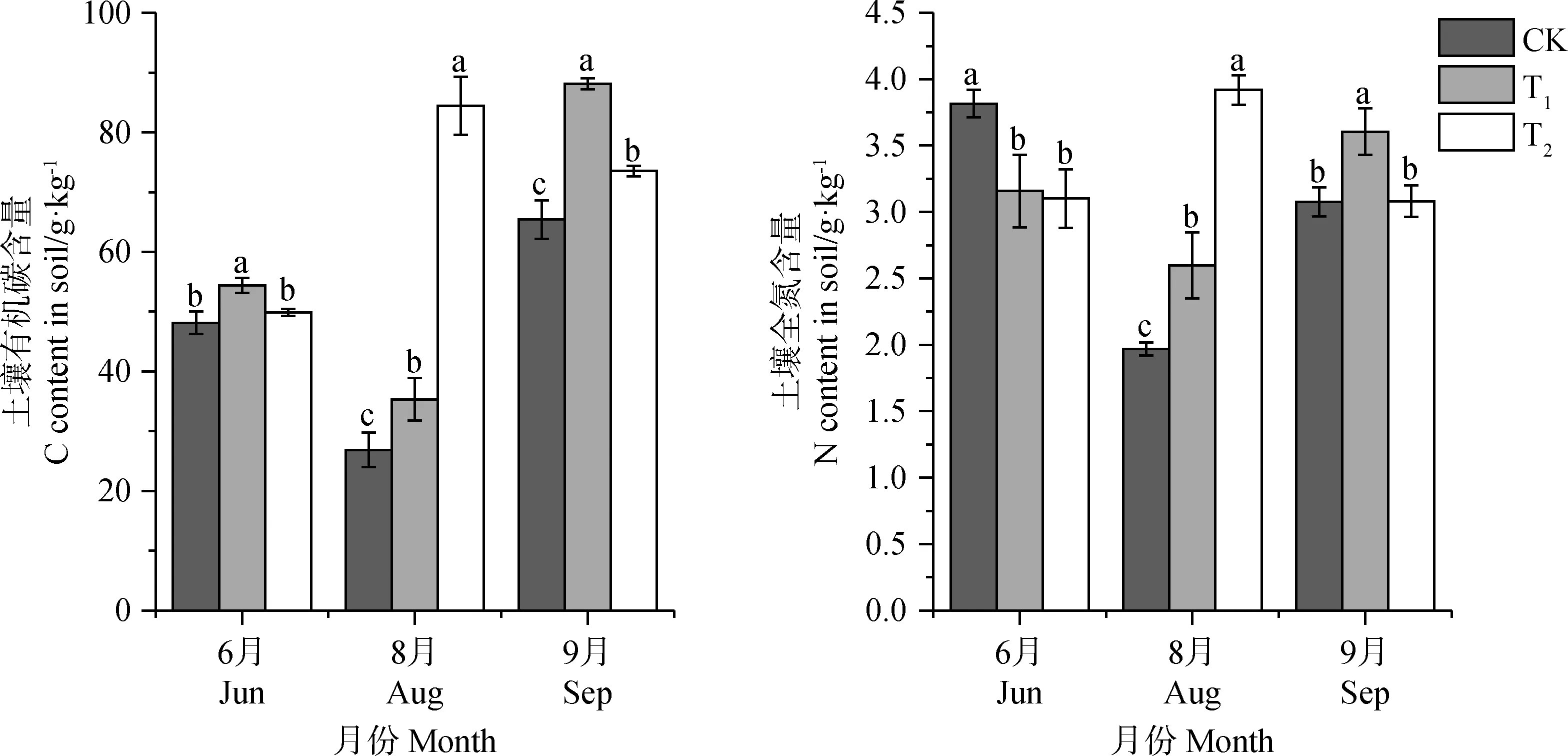
图5 不同增温幅度对土壤有机碳、全氮含量的影响。Fig.5 The effect of different temperature increase ranges on soil organic carbon and total nitrogen content in different seasons
2.6 增温处理下植物器官及土壤C/N的比较
增温处理对6月植物地上部分C/N表现为显著的促进作用(P<0.05),但8月显著低于对照(P<0.05),9月有一定的促进作用但差异并不显著。植物枯落物C/N明显增加,但其中8月T2处理及9月T1处理下差异并不显著,T1处理组相比CK增加了1.6%~14.5%,T2处理组相比CK增加了0.5%~36.7%。而对于增温后的植物根系C/N来说,除6月T1处理下差异并不显著外(P>0.05),其他不同时期、不同处理下的植物根系C/N显著降低(P<0.05)。T1处理下根系C/N降低了8.1%~12.2%,T2处理组相比CK使根系C/N降低了13.9%~28.8%,表明增温处理减少了植物根系C/N。表层土壤C/N与之相反,除8月T1处理下差异并不显著外(P>0.05),其他不同时期、不同处理下的土壤C/N明显增加。T1处理组相比CK增加了0.8%~37.6%,T2处理组相比CK增加了14.4%~57.8%。说明在整个生长季内增温明显影响了植物对氮素的利用、枯落物的分解以及土壤C,N供给和储存(表1)。

表1 不同处理对各组分C/N的影响Table 1 Effect of different treatments on C/N of each component
注:不同小写字母表示同一指标在同一时间下不同处理差异显著(P<0.05)
Note:Different lowercase letters indicate that the same index differs significantly at the same time at the 0.05 level
同时,对不同增温处理下的植物与土壤C,N及其化学计量比之间的相关性分析得出:植物地上部分全氮和植物根系有机碳、植物根系C/N,植物地上部分C/N和土壤C/N,根系有机碳和枯落物全氮,根系C/N和枯落物全氮,土壤有机碳和土壤全氮均存在显著的正相关关系(P<0.01);植物地上部分全氮和枯落物全氮、土壤C/N,植物根系有机碳和枯落物C/N、土壤有机碳、土壤C/N,植物根系C/N和土壤有机碳、土壤C/N均存在显著的负相关关系(P<0.01);枯落物有机碳和植物根系有机碳、根系C/N间均存在显著的正相关关系(P<0.05);植物地上部分有机碳和植物地上部分全氮、植物根系全氮,植物地上部分C/N和枯落物全氮、植物根系有机碳、植物根系C/N,枯落物有机碳和土壤有机碳、土壤C/N,植物根系C/N和土壤全氮,土壤C/N和枯落物全氮之间均存在显著的负相关关系(P<0.05)(表2)。
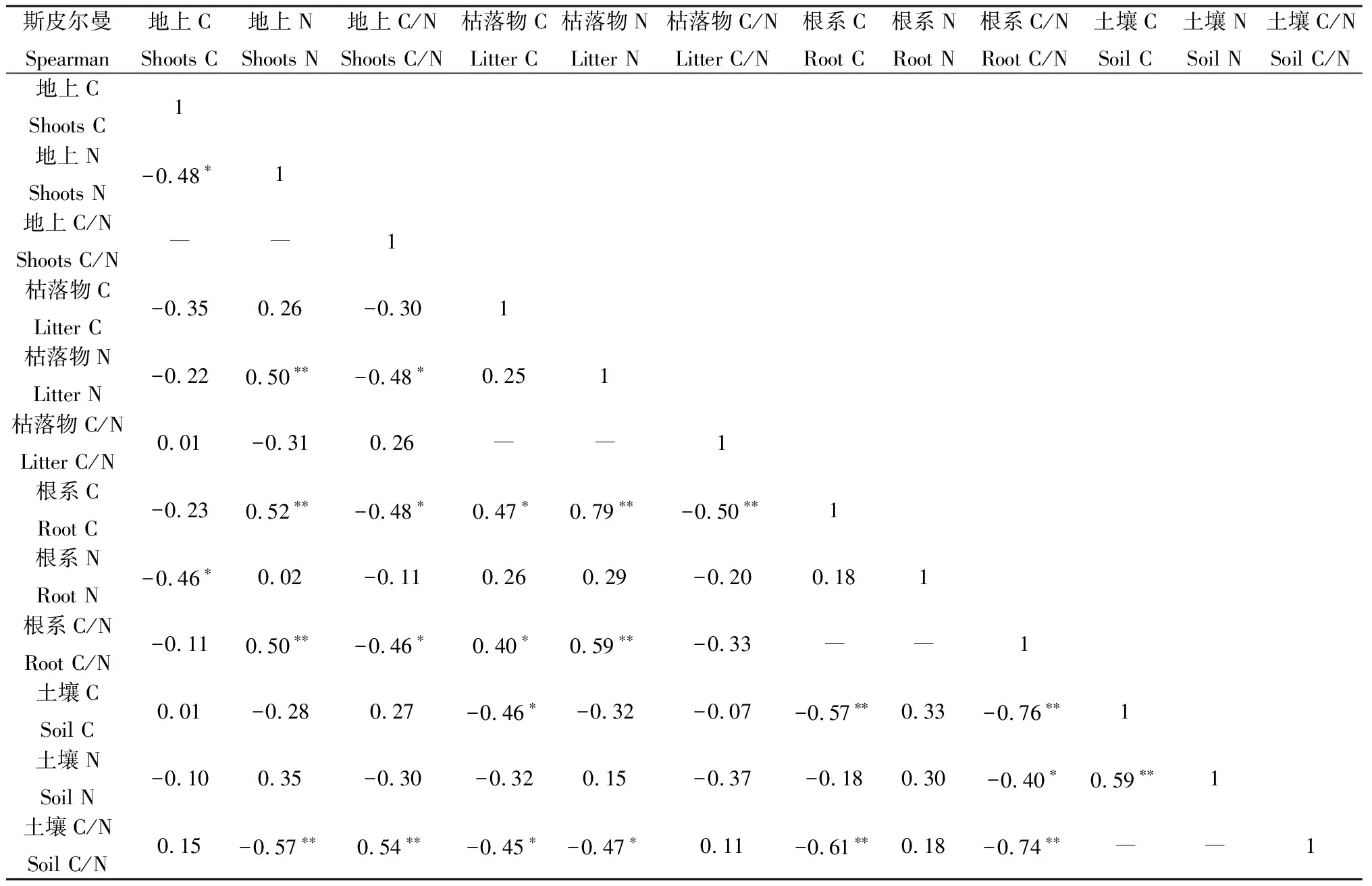
表2 植物、枯落物、土壤间C,N的相关性Table 2 The correlations of plants,litter and soil C and N
注:*,**分别代表显著性水平小于0.05和0.01;“―”表示存在自相关关系,不宜进行相关分析
Note:*,** represent the significance difference at the 0.05 and 0.01,respectively;"―" indicates that there is an autocorrelation relationship,and it is not appropriate to conduct correlation analysis
3 讨论
3.1 模拟增温对植物C,N的影响
本研究发现在整个生长季内植物地上部分碳含量除结实期T2处理下显著降低外,其他不同时期处理下并无显著变化,而根系碳含量除返青期外显著减少。这可能是由于植物在生长旺盛期,为了吸收更多的水分和养分,大量的碳水化合物被输送到根部以促进根系的生长,造成其根部碳水化合物的积累小于根系生长,从而导致根系C浓度显著降低。与本研究结果不同的是,Yang[16]等通过模拟增温对青藏高原植物养分分配的影响试验中发现,增温和干旱会降低植物地上部分C含量,增加植物根系C含量;而石福孙[17]等对川西北高寒草甸进行模拟增温试验却发现增温对植物地上部分C含量影响并不显著,使其根系C含量明显增加。由此可见,植物C含量对增温的响应并不一致,原因可能与植被类型的不同、所处季节的不同、增温方式、增温时间以及环境因素等有着密切联系。
本研究中返青期植物地上部分N浓度随温度升高而降低,结实期N浓度随温度升高而增加,枯黄期N浓度降低但差异不显著。可能是由于增温环境下,植物的生物量和氮利用效率均有所提高,从而降低了植物组织的N浓度[18-19];同时,由于结实期温度升高,光合作用增强,植物生长迅速,有机质积累量增加,增温加剧了微生物从有机物质中分解氮的能力[20],植物对土壤中可利用氮的吸引也随之增加,从而导致结实期地上活体N浓度随温度升高而增加。
地下根系N浓度与地上不同,T1处理下除结实期显著降低外(P<0.05),其他时间并无明显变化;在T2处理下返青期、结实期均显著增加(P<0.05)。可能是因为7,8月环境温度达到最大值,增温处理下温度超过了植物生长的适宜温度,使其处于半干旱条件下,植物除了进行光合作用,还需要增加氮和磷的吸收量,用以增加自身抗旱能力[21]。低增温处理下,处于8月结实期的植物根系向更深生长,以便吸收所需要的水分和营养物质带来了稀释效应,植物根对营养元素的吸收赶不上细胞膨胀的速率,使得根系N元素降低[22];而高增温处理下,表土的过于干旱对根的伸长产生强烈的机械阻碍作用[23],因此其N浓度高于对照;Yang等[16]研究也发现由于可溶性蛋白质的积累,干旱条件下植物N浓度显著增加。
此外,植物叶片C/N的大小可以表示植物吸收单位养分元素含量所同化C的能力,在一定程度上反映了植物体内养分元素的利用率[6]。有研究表明,增温通过增加植物生产能力以及植物的生物量来影响它们对营养的吸收,会降低植物中的C/N[24]。还有一些研究则表明,在气候寒冷的地区,增温通过增加生物量的生产,反而会使C/N有所增加[25]。Yang等[26]对青藏高原高山篙草的研究表明,增温降低了根部的C/N,提高了植物叶片的C/N,这意味着植物可能会以提高N的利用效率来应对未来的气候变暖。本研究发现增温降低了不同生长季植物根系C/N,且除结实期植物地上部分C/N低于CK外,其他不同时期、不同处理的植物地上部分C/N均高于对照,表明增温条件下返青期、枯黄期植物能够更有效地利用N素,增加其利用效率。结实期植物地上部分C/N低于CK可能是由于8月处于生长旺季,增温处理下温度升高,光合作用增强,植物生长迅速,有机质积累量增加,微生物从有机物质中分解氮的能力加剧,对土壤中可利用氮的吸引增强,从而导致地上部分C/N降低。增温处理后植物地上部分、根系C/N均发生改变,说明温度升高将会导致植株C/N改变,从而影响其对养分的利用效率。
3.2 模拟增温对枯落物C,N的影响
植物在自身的生长过程中,通过光合作用产生有机物,并通过枯落物的分解以及根系的分泌作用将养分归还于土壤中[27]。因此,枯落物及其分解过程是联系植被和土壤的纽带,它对土壤植物养分自身的需求和调节起着关键的作用[28],而微生物的分解活动与待分解物的C/N密切相关[29]。有研究表明,当枯落物N含量升高或C/N值降低会加快枯落物的分解速率,加速养分循环[30]。本研究发现,不同季节的植物枯落物有机碳对增温的影响并不一致,增温增加了返青期植物枯落物有机碳含量,减少了结实期、枯黄期枯落物有机碳含量,枯落物全氮含量在整个生长季内明显降低,枯落物C/N明显增加,且不同增温幅度对其影响并不一致,说明植物氮素在不同季节不同增温条件下发生了不同程度的释放和迁移,温度升高将会导致枯落物C/N改变,从而影响沼泽草甸生态系统的分解速率。安卓等[31]研究发现,在环境中养分供应短缺时,植物以提高养分重吸收率为其养分利用的主导方式适应环境胁迫,这恰好与本研究结果相互印证。
3.3 模拟增温对土壤C、N的影响
土壤是草地生态系统的重要组成部分,为植物提供必需的营养和水分,是各种物质能量的转化场所[32],其养分含量的高低影响着植被生物量生产及其分配[33]。本研究表明,温度升高增加了整个生长季0~10 cm土壤表层中的有机碳含量,可能是因为温度升高虽然加快了土壤有机碳的分解速率[34],但同时温度升高也增加了植物生产力,地表植物枯落物、根系以及根系分泌物补充了土壤中的有机质[35],从而使得土壤有机碳含量增加。
就增温处理后的表层土壤全氮来看,如图8所示,增温降低了返青期0~10 cm土壤表层中的全氮含量,增加了结实期、枯黄期0~10 cm土壤表层中的全氮含量。这可能是由于返青期植物生物量小,增温促进植被生长,需要大量的蛋白质和核酸来满足植物逐渐增加的生长速率,从而吸收利用大量的土壤氮素[23],导致土壤氮的减少。而土壤氮素在夏季则以氮素固持为主[36],使得土壤表层全氮含量增加。亓伟伟[37]等研究也表明,短期温度增加有利于有机质分解,使更多有机氮分解为植物可利用的矿化氮,促进土壤表层全氮含量增加。
研究还发现增温明显增加了整个生长季0~10 cm土壤表层中的C/N,使之从12.63~20.87增加到13.76~24.50,且对照和增温处理下的土壤C/N均明显高于全国范围(11.9±0.4)内C/N值的平均值[38],可以发现风火山地区高寒沼泽草甸土壤在整个生长季内均处于氮素限制状态,增温促进植物生长,加剧了这种限制。
3.4 植物—土壤C,N化学计量比之间的关系
土壤作为植物养分的主要来源[33],对于植物C,N含量具有十分重要的影响;植物枯落物及其分解速率又反过来影响土壤C,N含量。通过分析植物地上部分、枯落物、根系以及土壤C,N化学计量比之间的相关性发现,枯落物C,N含量与植物地上部分相关,土壤养分含量受枯落物限制,根系C,N含量又与土壤养分含量有关,植物地上部分C、N含量又与根系C、N含量有关,生态系统内部C元素和N元素在植物地上部分、枯落物、根系与土壤之间实现了运输和转换。植物根系从土壤中吸收养分供植物生长,地上部分通过光合作用合成有机物质,又以枯落物分解的形式将养分归还于土壤或损失[39],从而使得不同季节不同增温处理下C元素和N元素地上部分>枯落物>根系>土壤。
4 结论
通过在青藏高原腹地风火山地区高寒沼泽草甸开展的模拟增温试验可知,无论地上植物、枯落物还是地下根系,其C,N含量与土壤C,N含量之间均存在显著相关(P<0.05)或极显著相关(P<0.01);同时发现该研究区土壤处于氮限制状态(C/N>12),整个生长季内增温和对照处理下的植物生长均受N素限制,增温在一定程度上促进植物生长,加剧了该研究区氮素的缺乏状况,从而导致植物对C,N的利用率和枯落物的分解速率改变,土壤C/N明显增加。
猜你喜欢
现代园艺(2022年7期)2022-11-19
四川劳动保障(2021年3期)2021-06-09
落叶果树(2021年6期)2021-02-12
文苑(2020年12期)2020-04-13
海峡姐妹(2019年1期)2019-03-23
现代园艺(2017年13期)2018-01-19
现代农业(2016年3期)2016-04-14
中国农业文摘-农业工程(2016年5期)2016-04-12
土壤与作物(2015年3期)2015-12-08
郑州大学学报(理学版)(2012年4期)2012-03-25
