
苜蓿SKU5基因的表达调控及其在秋眠中的作用
2019-12-20 01:27杜红旗徐照学房卫平冯长松娄治国袁华祎
草地学报 2019年6期
杜红旗, 徐照学, 房卫平, 冯长松*, 娄治国, 袁华祎
(1.河南省农业科学院畜牧兽医研究所, 河南 郑州, 450008; 2.河南省畜禽繁育与营养调控重点实验室, 河南 郑州, 450008;3.河南省农业科学院经济作物研究所, 河南 郑州, 450008)
紫花苜蓿(MedicagosativaL.)是世界上广泛种植和利用的多年生优良豆科牧草,在其栽培和品种选育中考虑的首要指标是秋眠性。秋眠性是指夏末秋季由于日照长度缩短和温度下降,紫花苜蓿植株表现为缓慢匍匐生长或停止生长的一种特性。根据苜蓿秋眠特定时期刈割后植株的再生高度,将其分为秋眠型苜蓿(秋眠1-3级)、半秋眠型苜蓿(秋眠4-6级)和非秋眠型苜蓿(秋眠7-9级)[1-2],后来又扩展到 11 个秋眠级别(10-11 级),包括 2个极不秋眠苜蓿[3]。秋季时秋眠型苜蓿品种生长缓慢、叶小和茎纤细,但具有较强的抗寒性和高的越冬率;相反,非秋眠型苜蓿依然生长旺盛,但抗寒性差[4-5];半秋眠型苜蓿介于2者之间。
目前,国内外普遍认为调控紫花苜蓿秋眠性的主要环境因子是光周期和温度[6-7],其中光周期是诱导苜蓿秋眠的关键因子[8]。国外对不同季节不同秋眠类型苜蓿根系的生长状态、根系和根颈可溶性糖的积累,以及根和芽氮素化合物的分解代谢进行了大量研究[9-13],并且2000年Douglas等人确定了控制秋眠型苜蓿秋季生长的基因组区域,但是该区域具体的核苷酸序列未知[14]。国内对苜蓿品种的遗传多样性、秋眠性与抗寒性关系、秋眠性与生产性能的关系等方面进行了较多的研究[15-17],认为光敏色素、隐花色素2B、脱落酸,生长素等可能参与秋眠性的调控[18-20]。王成章、杜红旗等应用高通量测序技术得到了大量的调控苜蓿秋眠的候选基因、蛋白和microRNAs,并且选出了重要候选调控基因和蛋白[21-24]。但是这些仅仅停留在调控秋眠性候选基因和蛋白的筛选阶段,并未真正确定调控苜蓿秋眠性的基因。
前期应用转录组测序筛选调控苜蓿秋眠性关键基因时发现单铜氧化酶样蛋白SKU5基因(SKU5)在秋眠型苜蓿Maverick叶片中秋眠时期的表达量显著高于未秋眠时的表达量[23],因此推测它可能调控苜蓿的秋眠性。单铜氧化酶样蛋白SKU5属于植物多铜氧化酶亚家族成员SKS蛋白,SKS家族蛋白是植物多铜氧化酶基因家族中存在的一类特别的亚家族,均属于定位在细胞质膜上的糖基化蛋白。它们只包含多铜氧化酶中保守的T2Cu连接位点而缺乏其他类型铜离子连接所需的组氨酸残基,其可能已经不具备正常的多铜氧化酶功能,而是以一种新的方式作用于植物生长发育,但是这一领域的认识几乎是空白,目前唯一确定的是,这种变异并不是偶然的,对于植物机体来说,这种新型的多铜氧化酶一定有着不可取代的生理功能,这些都需要更加深入探索[25]。目前,在拟南芥中发现了SKS蛋白家族单铜氧化酶样蛋白SKU5和SKS1-SKS18共19个成员,在油菜、烟草、玉米、报春花等植物中也发现了该类蛋白。SKS家族基因编码的蛋白质在植物根生长方向、叶脉形式、花粉发育、花粉管伸长、激素调节等生物学过程中均具有作用,但是整个SKS基因家族的研究只集中在少数基因上[26-31]。
Sedbrook等最先从拟南芥(Arabidopsisthaliana)中得到了SKS家族成员基因SKU5。SKU5全长为3.3Kb,包含9个外显子,开放阅读框长1 764 bp,预期编码一个定位在细胞质膜和细胞壁上的大小为66kDa的糖蛋白,其结构类似于多铜氧化酶中的AAO和漆酶。虽然单铜氧化酶样蛋白SKU5与这2种酶相似,但是它缺乏T1Cu和T3Cu所需的组氨酸残基。SKU5在拟南芥各组织中均有表达,而在根尖分生组织和末梢伸长组织中表达量最大。SKU5过表达植株在琼脂糖上生长时,根和下胚轴与野生型相比较短15%,根偏离了正常的向下生长,并且与黄化的下胚轴都显示出逆时针的轴向旋转,打乱了根系的正常向下生长[27]。但是目前苜蓿SKU5的研究未见报道。
本研究通过检测春夏秋季和人工条件下不同光照时间或温度下秋眠型苜蓿品种Maverick和非秋眠型苜蓿品种Cuf101叶片中SKU5 mRNA相对表达量及分析其与苜蓿株高、叶面积、日照长度、温度的相关性来研究它在苜蓿秋眠中的作用。本研究为阐明苜蓿秋眠性分子机理提供了新的内容、丰富了苜蓿秋眠性理论,也为苜蓿分子育种提供功能基因。
1 材料与方法
1.1 样品采集、株高和叶面积检测并记录日照长度和温度
试验材料为美国苜蓿标准秋眠1级品种Maverick和秋眠9级品种Cuf101。2011年4月份到10月份期间测株高、叶面积,记录日照长度、温度并取样。具体操作是:每次刈割后第14天上午8:00采集各品种至少三株苜蓿植株的叶片,取每个分枝顶端第一个成熟叶片以下的第3—5片成熟叶片,取后立刻保存在液氮中,之后存于-80℃冰箱待用,同时记录每个月的日照长度和温度。每一苜蓿品种随机选取5株,每株随机选取10片叶片测长和宽,应用A=KLW (A:叶面积leave area;K:修正系数coefficient of correction;L:叶长Leave length;W:叶宽leave wide)公式计算叶面积[32]。每次取样时每一苜蓿品种随机选取15株,每株测高、中、低枝条高度取平均值作为该苜蓿株的株高。
Maverick和Cuf101苜蓿在人工气候箱中不同光照时间或不同温度下培养处理。每种苜蓿各3组,每组各3株苜蓿植株,分别在每天8 h,12 h和16 h的光照时间培养,温度均为24℃、光照强度均为3 000 lux。每种苜蓿另外各3组,每组各3株苜蓿植株,分别在16℃,24℃和 32℃下培养,光照强度均为3 000 lux、每天光照时间均为16 h。每种处理均是刈割后开始处理,处理14d后取各自的叶片并储存于-80℃冰箱待用。
1.2 总RNA提取与反转录
严格按照TRizol法提取总RNA的操作步骤提取各样品的总RNA(Invitrogen,Carlsbad,CA,USA),应用Nano2000核酸检测仪器(ThermoFisher Scientific,Waltham,MA,USA)检测各样品RNA的浓度,并调至同一浓度,严格按照TAKARA公司反转录试剂盒的说明书上操作步骤对各样品的总RNA反转录。
1.3 各样品中SKU5 mRNA相对表达量检测
前期转录组测序得到的SKU5部分序列在NCBI数据库中序列比对进行基因注释、保守序列分析和各外显子界限的分析,根据序列分析结果和紫花苜蓿GAPDH基因序列应用primer5.0软件按照荧光定量引物设计原则设计SKU5和GAPDH的荧光定量引物(SKU5-S:GGAACACCAAGAGGAGAAGCA,SKU5-A:TTTTCATCCTCAACATTGCTCTG;GAPDH-S:TGGGAAGCACATTACAGCAG,GAPDH-A:CATCAGCATTGACACCAACC),在罗氏Cycler9.0荧光PCR仪上应用罗氏SYBER green荧光染料检测各样品中SKU5mRNA相对表达量。
1.4 统计学分析
采用2-ΔΔCt公式计算SKU5mRNA相对表达量,用平均值±标准差表示[33]。应用SPSS 19.0(IBM Corp.,USA)软件对所测数据进行统计学分析。应用one-way ANOVA方法分析各样品间SKU5 mRNA相对表达量的差异显著性,并应用双因子相关性分析和双边T检验方法分析SKU5 mRNA相对表达量、日照长度、温度、株高和叶面积之间的相关性。最后,应用GraphPad prism 5 (GraphPad Software,Inc.,USA)软件对各数据进行线型图的制作。
2 结果与分析
2.1 采样时期日照长度和温度
从4月份至10月份,日照长度和温度的变化趋势均先升高后降低。从4月份至6月份的夏至,日照长度逐渐延长,其中夏至日最长;从6月份夏至日始至10月份,日照长度逐渐缩短;从4月份至10月份,温度先升高后逐渐下降,其中7月份最高(图1)。
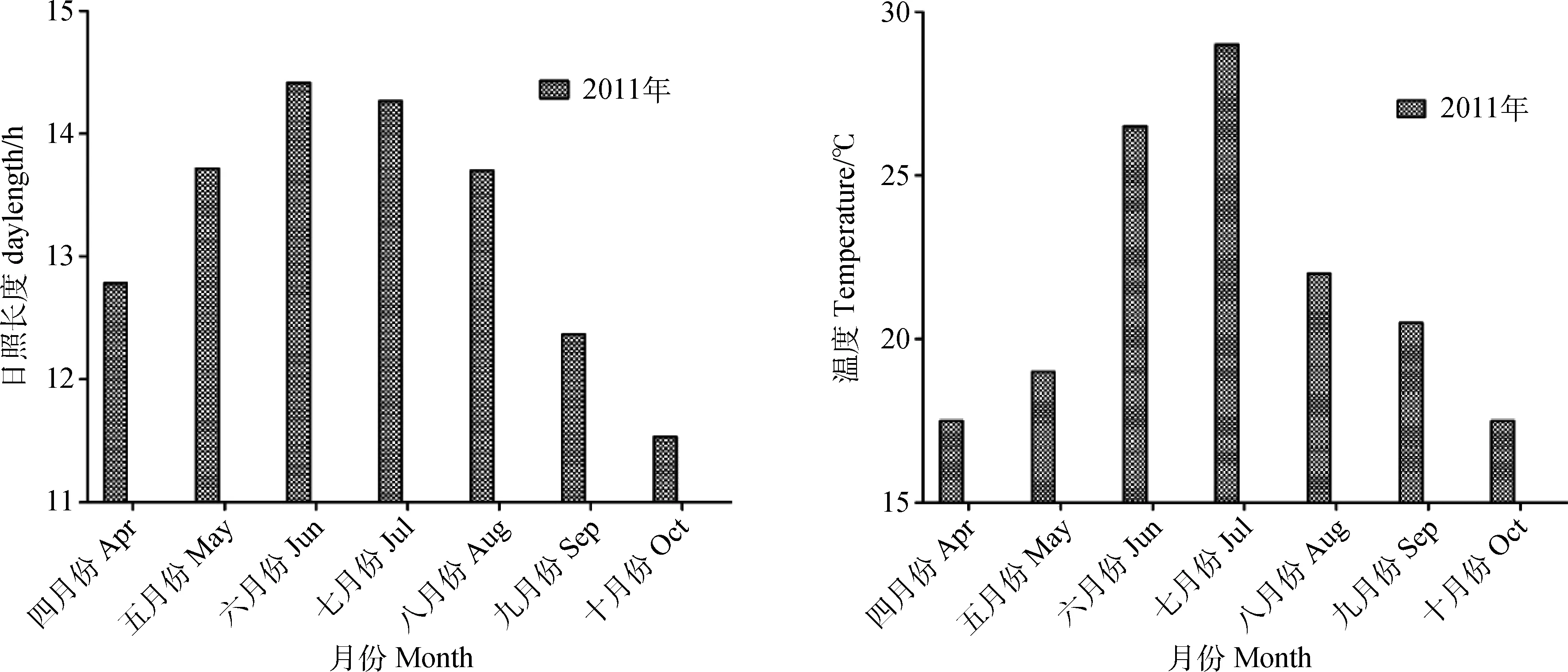
图1 4—10月份的日照长度和平均温度Fig.1 Daylength and average temperature from April to October
2.2 采样时2种苜蓿株高和叶面积
5月份至10月份,Maverick品种的株高逐渐下降,4—8月份Maverick苜蓿株高两两差异不显著,9,10月份株高极显著低于4—8月份的株高(P< 0.01),表明Maverick品种苜蓿在秋季发生秋眠;Cuf101株高4—10月份之间差异不显著,但6—9月份株高有下降的趋势,7—10月份Cuf101苜蓿株高两两月份之间差异不显著;但是Cuf101苜蓿的株高显著高于对应月份Maverick苜蓿的株高(P<0.05),表明Cuf101苜蓿在秋季不发生秋眠(图2)。
Maverick苜蓿叶面积小于Cuf101苜蓿叶面积。8,9和10月份Maverick苜蓿叶面积极显著小于Cuf101苜蓿叶面积(P< 0.01);8,9和10月份Maverick苜蓿叶面积显著小于其它时期叶面积(P<0.05);Cuf101苜蓿叶面积各月份之间差异不显著,表明秋季Maverick苜蓿秋眠时的叶面积显著变小(图3)。
2.3 SKU5基因mRNA相对表达量
2.3.1自然条件下SKU5基因mRNA相对表达量 自然条件下,从4月份至10月份秋眠型苜蓿Maverick叶片中SKU5 mRNA相对表达量呈先升高后降低的趋势;非秋眠型苜蓿Cuf101叶中SKU5 mRNA相对表达量呈升高、降低、升高、降低的趋势;秋眠型苜蓿Maverick叶片中SKU5 mRNA相对表达量在9月份最高,秋季时的相对表达量显著高于春夏季时的相对表达量(P<0.05);非秋眠型苜蓿Cuf101叶片中SKU5 mRNA相对表达量在8月份最高,之后SKU5 mRNA相对表达量急剧降低(图4)。结果表明:秋季时2种苜蓿叶中SKU5 mRNA相对表达量变化存在差异;秋眠型苜蓿Maverick叶中SKU5 mRNA相对表达量在秋眠时期的量高于未秋眠时期的量和秋季时非秋眠型Cuf101叶片中的量。
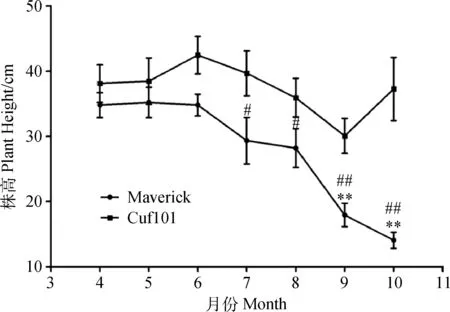
图2 4—10月份2种苜蓿株高Fig.2 Plant height of Maverick and Cuf101 varieties from April to October注:**代表Maverick苜蓿9,10月份株高极显著低于4月份时的株高(P < 0.01);#代表7,8月份Maverick苜蓿株高显著低于Cuf101株高(P< 0.05),##代表9,10月份Maverick苜蓿株高极显著低于Cuf101株高(P < 0.01)Note:** indicates that plant height of Maverick at September and October is very significant lower than that at April at the 0.01 level;# indicates that plant height of Maverick is significant lower than that of Cuf101 at July and August at the 0.05 level;## indicates that plant height of Maverick is very significant lower than that of Cuf101 at September and October at the 0.01 level

图3 两品种苜蓿4—10月份叶面积Fig.3 Leaf area of Maverick and Cuf101 varieties from April to October注:##代表8、9、10月份Maverick苜蓿叶面积极显著小于8,9,10月份Cuf101的叶面积(P<0.01),**代表9、10月份Maverick苜蓿叶面积极显著小于其4-7月份时的叶面积(P<0.01),*代表8月份 Maverick苜蓿叶面积显著小于其4-7月份时的叶面积(P<0.05)Note: ## indicates that leaf area of Maverick is very significant smaller than that of Cuf101 at August, September and October at the 0.01 level, ** indicates that leaf area of Maverick at September and October is very significant smaller than that at April to July at the 0.01 level, * indicates that leaf area of Maverick at August is significant smaller than that at April to July at the 0.05 level
2.3.2人工气候条件下SKU5基因mRNA相对表达量 人工气候条件下,随着日照长度延长,2种苜蓿叶片中SKU5 mRNA相对表达量逐渐减少,其中8h日照长度时的量显著高于12h和16h时的量(P< 0.05);秋眠型苜蓿品种Maverick叶片中的量极显著高于非秋眠型苜蓿品种Cuf101叶片中的量(P< 0.01)(图5)。
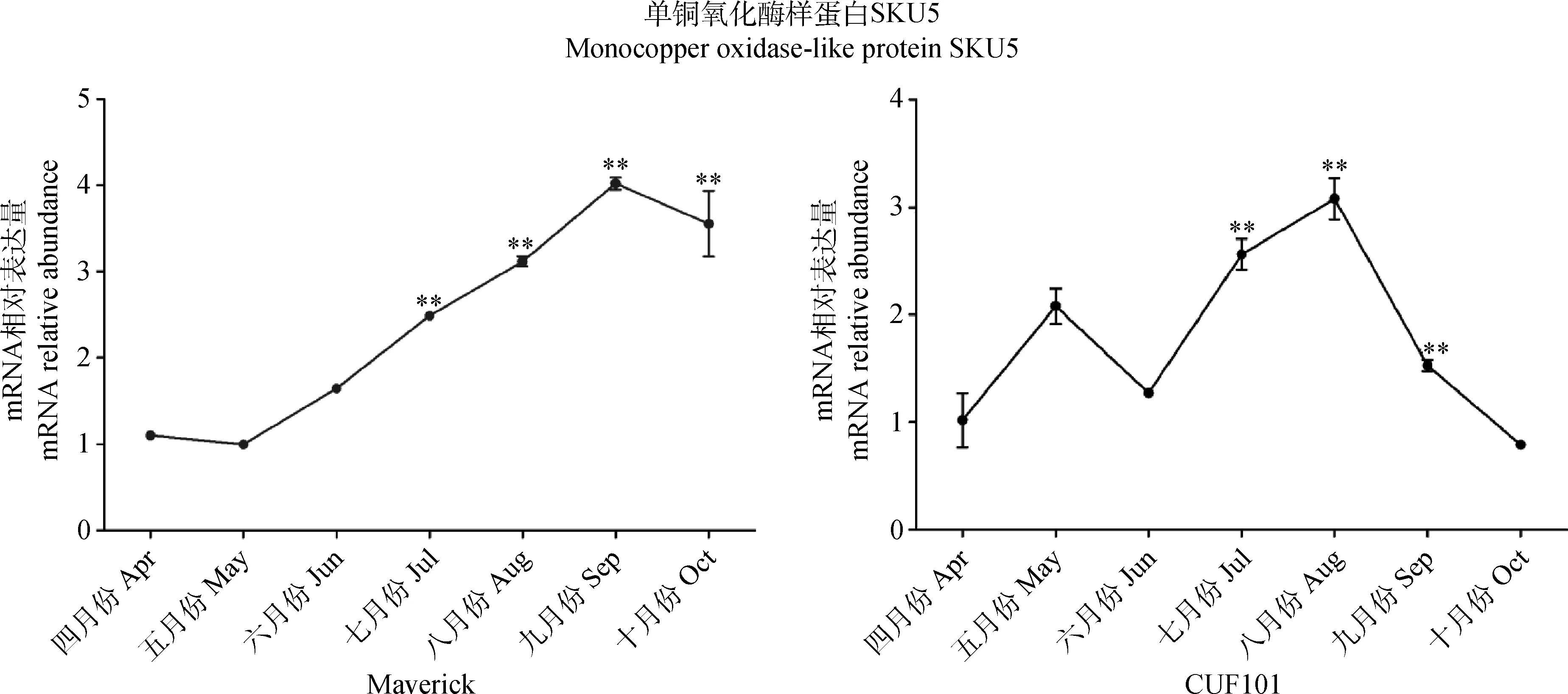
图4 4—10月份2种苜蓿叶中SKU5 mRNA相对表达量Fig.4 mRNA relative abundance of SKU5 in Maverick and Cuf101 varieties from April to October注:**代表7,8,9,10月份时的SKU5 mRNA相对表达量极显著高于4月份时的相对表达量Note:** indicate that SKU5 mRNA relative abundance in two varieties alfalfa at July,August,September,October is very significant more than that at April at the 0.01 level

图5 不同日照长度下两品种苜蓿真叶中SKU5mRNA相对表达量Fig.5 mRNA relative abundance of SKU5 in Maverick and Cuf101 varieties grown under 8h to 16h daylength and artificial growth conditions注:* 代表8 h下每一品种苜蓿叶片SKU5 mRNA相对表达量显著高于12 h和16 h下的相对表达量;##代表Maverick苜蓿SKU5 mRNA相对表达量极显著高于Cuf101苜蓿中的相对表达量Note:* indicates that the mRNA relative abundance of SKU5 in Maverick and Cuf101 varieties under 8 h illumination is significant more than that 12 h,and 16 h of illumination at the 0.05 level;## indicate that the mRNA relative abundance of SKU5 in Maverick leaves is very significant more than that in Cuf101 leaves at the 0.01 level
随着温度的升高,苜蓿叶片中SKU5 mRNA相对表达量逐渐增加,其中16℃时的量极显著低于24℃和32℃时的量(P< 0.01),Maverick叶片中的量极显著高于Cuf101叶片中的量(P< 0.01)(图6)。结果表明:人工气候条件下,秋眠型苜蓿Maverick和非秋眠型苜蓿Cuf101叶片中SKU5的表达受光周期和温度的调控规律一致,但是2种苜蓿品种叶片中SKU5基因的转录受日照长度和温度调控趋势相反。
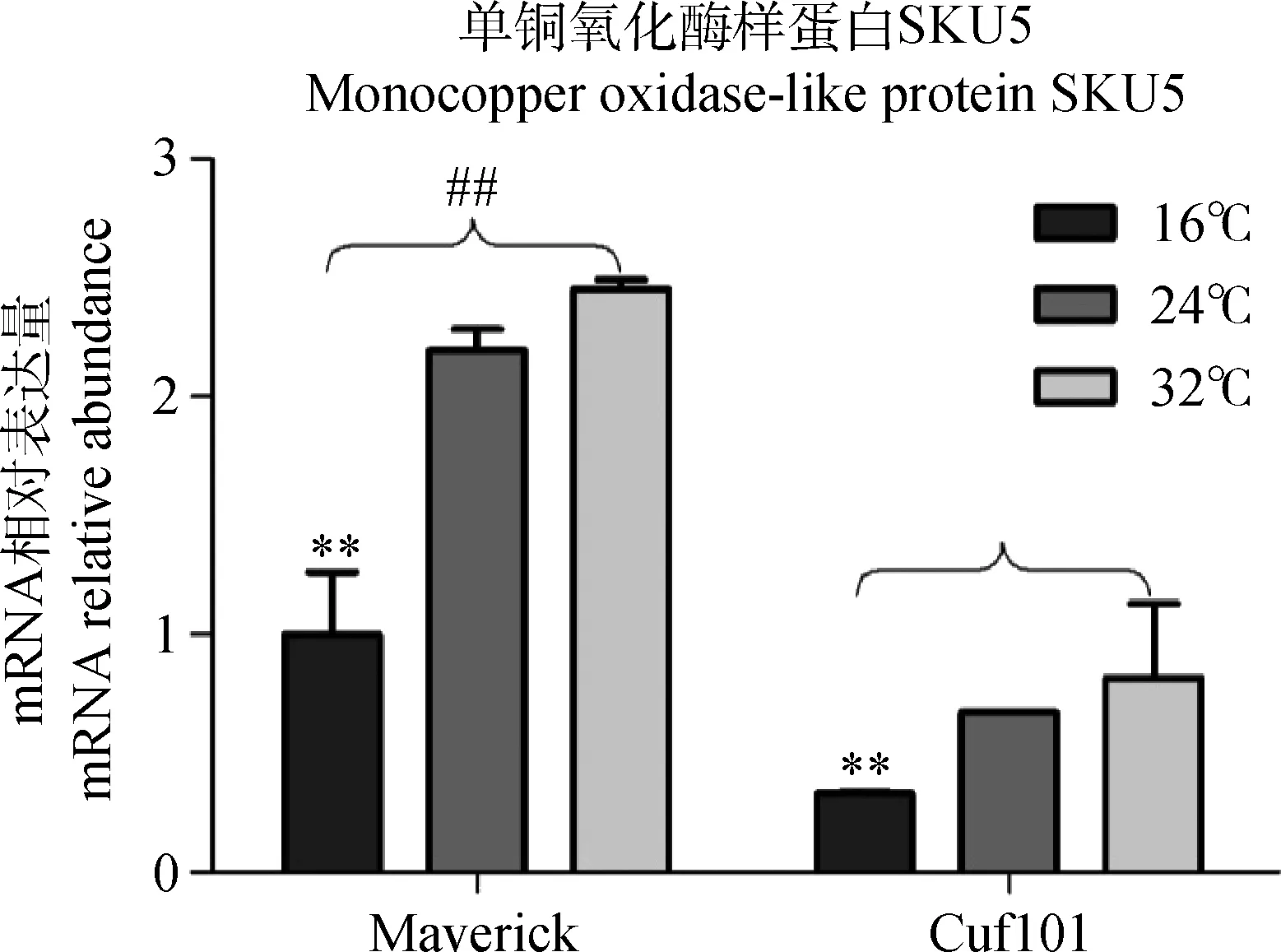
图6 人工条件下不同温度处理的两品种苜蓿叶中SKU5 mRNA相对表达量Fig.6 mRNA abundance of SKU5 in Maverick and Cuf101 varieties grown under artificial growth conditions at different temperatures (from 16℃ to32℃)注:** 代表16℃下每一品种苜蓿叶片SKU5 mRNA相对表达量极显著低于24℃和32℃下的相对表达量;##代表Maverick苜蓿SKU5 mRNA相对表达量极显著高于Cuf101苜蓿中的相对表达量Note:** indicate that the mRNA relative abundance of SKU5 in both varieties at 16℃ is very significant less than that 24℃and 32℃at the 0.01 level;## indicate that the mRNA relative abundance of SKU5 in Maverick leaves isvery significant more than that in Cuf101 leaves at 16℃,24℃ and 32℃ at the 0.01 level
2.4 SKU5mRNA相对表达量、苜蓿叶面积、株高、日照长度、温度之间的相关性
自然条件下,SKU5 mRNA相对表达量与日照长度、温度没有显著相关性(表1);但是在人工气候条件下,SKU5 mRNA相对表达量与日照长度极显著负相关、与温度极显著正相关;日照长度和温度对SKU5表达的调控趋势相反(表2)。
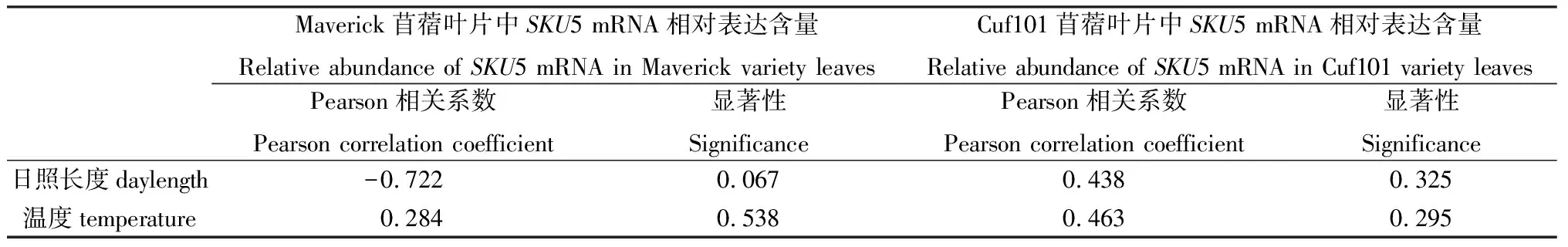
表1 自然条件下SKU5 mRNA相对表达量与日照长度、温度相关性Table 1 Correlation between SKU5 mRNA relative abundance in the leaves of two varieties and daylength,temperature under natural environment
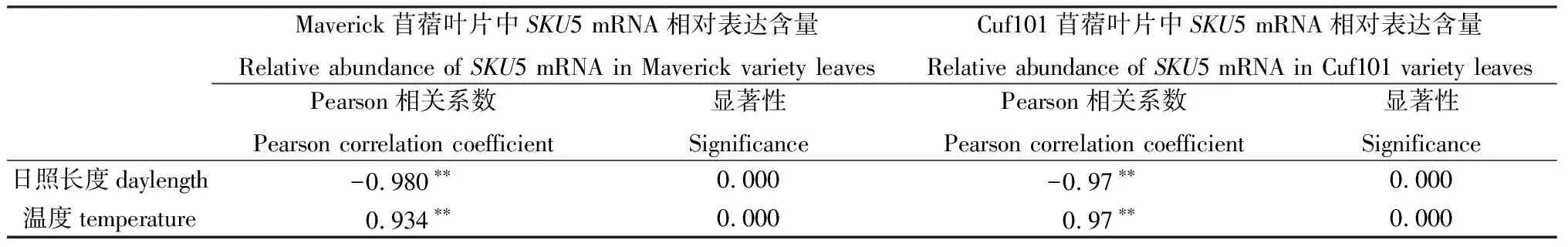
表2 人工条件下SKU5 mRNA相对表达量与日照长度、温度相关性Table 2 Correlation between SKU5 mRNA relative abundance in the leaves of two varieties and daylength,temperature under artificial growth conditions
注:** 代表差异极显著(P<0.01)
Note:** indicate significant correlation at the 0.01 level
自然条件下,日照长度、温度与2种苜蓿株高无显著相关性,但日照长度与2种苜蓿株高相关性更高,与秋眠型苜蓿Maverick株高的相关性和显著性高于与非秋眠型苜蓿Cuf101的相关性和显著性(表3)。
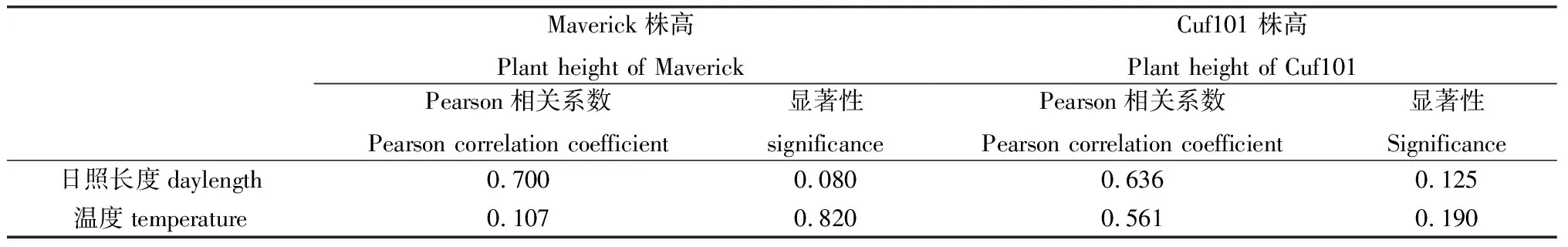
表3 日照长度、温度与2种苜蓿株高相关性Table 3 Correlation between daylength,temperature and the plant height of two varieties
自然条件下,秋眠型苜蓿Maverick叶中SKU5 mRNA相对表达量与叶面积、株高呈显著负相关;而非秋眠型苜蓿Cuf101叶中的相对表达量与叶面积、株高无显著相关(表4)。
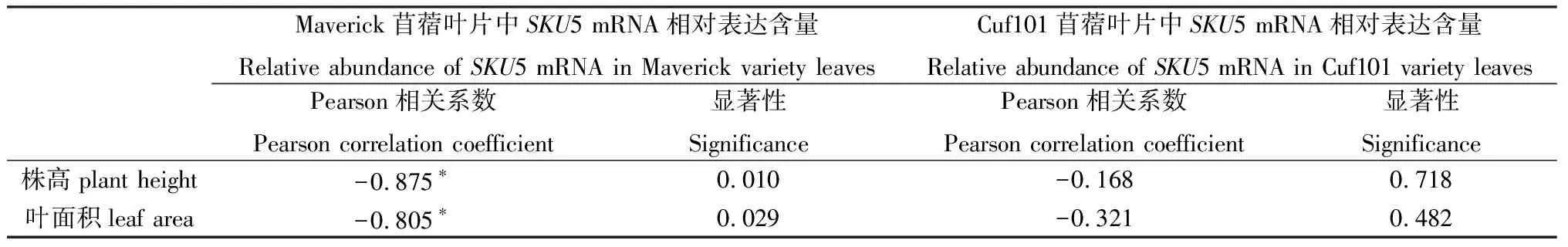
表4 SKU5 mRNA相对表达量与株高、叶面积相关性Table 4 Correlation between SKU5 mRNA relative abundance in the leaves of two varieties and plant height,leaf area
注:* 代表差异显著(P<0.05)
Note:* indicates significant correlation at the 0.05 level
以上结果表明:在日照长度与温度共同作用的自然条件下,随着秋眠型苜蓿Maverick叶片中的SKU5基因可能通过上调其表达抑制叶片和植株的生长;非秋眠型苜蓿Cuf101SKU5基因影响其叶片和植株的生长不显著。
3 讨论
不同秋眠型苜蓿在不同季节生长表现差异,特别是在秋季生长表现显著差异并用秋眠的程度来衡量,但是目前苜蓿秋眠的发生机理不清楚。本研究表明:秋眠型苜蓿Maverick和非秋眠型苜蓿Cuf101叶片中SKU5基因的表达受日照长度极显著负调控、受温度极显著正调控,但自然条件下,秋眠型苜蓿Maverick叶片中SKU5基因的表达主要受日照长度的调控,非秋眠型苜蓿Cuf101叶片中SKU5基因的表达主要受温度的调控;秋眠型苜蓿MaverickSKU5基因的高表达减小其叶面积、降低其株高从而促进其秋眠,而非秋眠型苜蓿Cuf101SKU5基因的表达高低对其叶面积和株高变化的影响不明显。
3.1 两种秋眠型苜蓿叶片中SKU5基因的表达受日照长度和温度调控能力不同
在人工条件下2种秋眠型苜蓿SKU5基因的表达受日照长度和温度调控的趋势均相反,日照长度负调控SKU5基因的表达(表1),日照长度的缩短促进SKU5基因的表达、温度的降低减少其表达;在自然条件下该基因的表达是受日照长度和温度双因子共同作用的结果,因此得不到自然条件下SKU5基因的表达受日照长度和温度的调控(图4)。对于秋眠型苜蓿Maverick,从6月份开始随着日照长度的缩短SKU5基因的相对表达量增加,从7月份起温度的降低并未改变SKU5基因表达增加的趋势,因此秋眠型苜蓿Maverick叶片中SKU5基因的表达主要受日照长度的调控。从非秋眠型苜蓿Cuf101叶片中SKU5基因的表达整体上看,高温时非秋眠型苜蓿Cuf101叶片中SKU5基因的相对表达量高于低温时的相对表达量,随着温度的降低其相对表达量减少,但从8月份,随着日照长度的缩短非秋眠型苜蓿Cuf101叶片中SKU5基因的相对表达量并未增加,因此非秋眠型苜蓿Cuf101叶片中SKU5基因的表达主要受温度的调控。秋眠型苜蓿最初引自寒冷气候地区、具有很好的抗寒性,适合于寒冷气候的地区生长;而非秋眠型苜蓿引自冬季温度温和的地区,适合于热带地区生长;它们形成了各自独有适应环境的调控方式[1],并且秋眠型苜蓿与非秋眠型苜蓿的遗传背景和遗传多样性不同[34-35]。因此,由于环境因子的变化,各自调控基因表达的调控机理和结果表现出大的差异,这可能就是导致本研究中两种秋眠型苜蓿叶片中SKU5基因的表达受日照长度和温度调控能力不同的原因。
在自然条件下,4—10月份2种苜蓿SKU5基因的表达趋势基本一致,但也有差别,最大的差别是秋季时二者SKU5基因mRNA相对表达量变化不一致,秋眠型苜蓿Maverick叶片中SKU5基因mRNA相对表达量在9月份达到最高,10月份缓慢减少,而非秋眠型苜蓿Cuf101叶片中SKU5基因mRNA相对表达量在8月份达到最高,9,10月份迅速降低(图4),在这个时期秋眠型苜蓿发生秋眠,非秋眠型苜蓿不发生秋眠。因此我们推测:2种苜蓿中SKU5基因表达的差异在秋季它们生长表现差异中起着重要的作用,在秋季它的高表达促进了秋眠型苜蓿的秋眠、低表达维持了非秋眠型苜蓿的正常生长。SKU5基因的过表达拟南芥相对于野生型植株其根和下胚轴变短[27],此结果与本研究的结果相似。
3.2 SKU5基因调控苜蓿秋眠性机理
叶面积是苜蓿秋眠性的一个重要指标[21,36]。它是叶片细胞周期、分裂和生长的宏观表现,细胞周期调节与整个器官生长速率的变化有着直接的联系,细胞周期、分裂和生长受细胞和环境因素之间空间关系的调节[37-38],叶片大小受细胞周期、分裂和生长相关基因表达变异的影响[39],有研究表明单铜氧化酶样蛋白SKU5与细胞周期、分裂和生长均有关[27]。
日照长度是调控苜蓿秋眠性的关键环境因子[16-17],从夏季至秋季随着日照长度的缩短秋眠型苜蓿发生秋眠。本研究表明SKU5基因的表达受日照长度和温度的调控,其中秋眠型苜蓿Maverick叶片中SKU5基因的表达主要受日照长度的调控,日照长度的缩短导致秋眠型苜蓿叶片中SKU5基因的表达量升高(图4),而且秋眠型苜蓿Maverick叶片中SKU5基因mRNA的相对表达量与其叶面积和株高显著负相关,SKU5基因的高表达减少了叶面积、降低了株高(图2-4,表4),秋季时秋眠型苜蓿发生秋眠。因此,随着日照长度的缩短,SKU5基因的高表达很可能促进了秋眠型苜蓿的秋眠。
众所周知,叶的光合作用、呼吸作用和蒸腾作用对植物的生长发育起着重要的作用。铜是蛋白质的重要辅助因子,参与呼吸代谢中的氧化还原反应,叶绿体质体蓝素中铜参与光合作用的电子传递过程[40],所以铜含量的减少会降低叶片光合作用效率。单铜氧化酶样蛋白SKU5基因的高表达会与叶绿体质体蓝素竞争铜而导致叶绿体质体蓝素能结合的铜相对减少从而降低光合作用,进而抑制了植物的生长。另外,有研究表明:单铜氧化酶样蛋白SKU5可能通过参与细胞壁的膨胀来参与2个方向的定向生长过程[27]。因此,SKU5调控秋眠型苜蓿秋眠的机理可能是随着日照长度的缩短导致了它的高表达从而降低了细胞壁的膨胀,抑制了叶片2个方向的生长而导致了叶片变小和变薄,以及降低了叶绿体质体蓝素结合的铜,最终共同减弱了叶片光合作用、呼吸作用和蒸腾作用,从而减慢了整个植株的生长而发生秋眠。
4 结论
苜蓿叶片中SKU5基因的表达受日照长度和温度的调控,但是2种秋眠型苜蓿叶片中SKU5基因的表达受日照长度和温度的调控方式不同;对于秋眠型苜蓿,随着日照长度的缩短SKU5高表达参与调控苜蓿的秋眠。
猜你喜欢
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
现代畜牧科技(2021年9期)2021-10-13
现代畜牧科技(2021年4期)2021-07-21
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
星星·散文诗(2017年2期)2017-07-05
中国三峡(2017年4期)2017-06-06
为了孩子(3~7岁)(2016年6期)2016-05-14
