
硒对金黄色葡萄球菌感染的奶牛乳腺上皮细胞Nod2/MAPK/mTORs信号通路中关键因子mRNA转录水平的影响
2019-11-22 03:07关立增刘俊俊韩照清毕崇亮
中国预防兽医学报 2019年9期
关立增,王 亨,刘俊俊,王 娟,韩照清,毕崇亮*
(1.临沂大学农林科学学院,山东临沂 276005;2.扬州大学兽医学院,江苏扬州225009;3.江苏省动物重要疫病与人兽共患病防控协调创新中心,江苏扬州225009)
奶牛乳房炎是奶牛养殖业中常见疾病之一。研究表明,金黄色葡萄球菌(Staphylococcus aureus)是引起奶牛乳房炎最常见的病原菌之一[1]。S.aureus可以进入宿主细胞,激活炎症相关信号通路,引发炎症反应。而奶牛乳腺上皮细胞(Bovine mammary epithelial cells,bMECs)是病原微生物进入动物乳腺的第一道防线[2-3]。当感染发生时,bMECs 可以表达大量的病原相关模式识别受体(Pattern recognition receptor,PRR),并快速识别病原微生物的病原相关分子模式(Pathogen-associated molecular patterns,PAMP),从而引起炎症反应和免疫应答反应[4]。
硒(Se)是动物体内最重要的微量元素之一,研究表明硒可通过调控信号通路的方式参与炎症和免疫调节等多种生物学过程[5-6],硒可以调节TLR2 信号通路并减轻bMECs 的炎症反应[7]。与TLR2 信号通路功能相似,Nod2 信号通路在胞内炎症反应过程中也发挥着至关重要的作用,本研究对硒在bMECs Nod2/MAPK/mTORs 信号通路中对关键因子mRNA转录水平的影响进行研究,探究硒调控S.aureus诱导bMECs 炎症反应的具体机制。
1 材料与方法
1.1 主要实验材料S.aureus(ATCC29213)由扬州大学兽医学院外科教研室惠赠;TRIzol 购自Invitrogen 公司;SYBR Green PCR mix、反转录试剂盒购自TaKaRa 公司。
1.2 实验分组试验分为对照(Con)组(bMECs)、模型(Mod)组(bMECs+S.aureus)和实验组。其中实验组又分 3 个亚剂量组,即 Low 组(bMECs+2 μmol/L Se+S.aureus)、Mid 组(bMECs+4 μmol/L Se+S.aureus)和 Hig 组(bMECs+8 μmol/L Se+S.aureus),每组设 3个重复。
1.3 S.aureus 感染bMECs 炎症模型的构建参照文献[7],采用机械剪碎配合II 型胶原酶消化法培养原代 bMECs,将 bMECs 以 106细胞 / 孔接种于 6 孔板中,当细胞超过80 %的汇合度时,实验组用含不同浓度硒(2 μmol/L、4 μmol/L 和 8 μmol/L)的培养基替换原培养基,继续培养12 h 后PBS 洗涤3 次,将S.aureus按MOI 1 加入6 孔板中,于感染后的不同时间点(6 h、8 h 和10 h)收集细胞,采用TRIzol法提取其RNA。同时按1.2 中分组设对照(Con)组(bMECs)、模型(Mod)组,并提取其RNA。
1.4 硒作用后 bMECs 关键因子 mRNA 的 qRTPCR 检测将1.3 中各组各时间点提取的总RNA利用反转录试剂盒反转录获得cDNA 后,分别以其为模板,采用荧光定量(qPCR)方法检测Nod2、RIP2、JNK、AKT、mTOR、IL-8、IL-10 和 β-actin的mRNA 的转录水平,引物序列见表1,同时设β-actin 为参考基因。反应体系总体积为20 μL,其中SYBR Green PCR mix 10 μL,上下游引物(10 μmol/L)各 0.8 μL,cDNA 模板 2 μL,ddH2O 6.4 μL。PCR反应条件为:95 ℃ 2 min;95 ℃ 10 s、60 ℃ 30 s、72 ℃ 30 s,30 个循环。
1.5 数据统计分析按2-ΔΔCt相对定量法计算各基因的表达量。利用SPSS13.0 软件进行数据分析,结果以 Mean±SEM 的形式表示。p<0.05 为显著性差异,p<0.01 为极显著性差异。采用Graph pad prism 6.0 软件绘图。

表1 引物序列Table 1 Primers used in this study
2 结 果
2.1 硒对Nod2 信号通路中关键因子Nod2 和RIP2 mRNA 转录水平的影响通过qPCR 方法对Nod2 信号通路中关键因子Nod2 和RIP2 的mRNA 转录水平进行检测。Nod2 mRNA 检测结果显示,在S.aureus感染6 h 后与对照组相比,模型组细胞中Nod2 mRNA的转录水平显著上升(p<0.05),而实验组中2 μmol/L、4 μmol/L 和 8 μmol/L 的硒均能显著抑制 bMECs 中该基因 mRNA 的转录水平(p<0.05);在S.aureus感染8 h 后,与对照组相比,模型组细胞中Nod2 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 的硒能极显著抑制bMECs 中该基因的转录水平(p<0.01),4 μmol/L 和 8 μmol/L 的硒能抑制该基因mRNA 的转录水平,但与模型组相比无统计学差异(p>0.05);在S.aureus感染 10 h 后,与对照组相比,模型组中Nod2 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 和 4 μmol/L的硒均能显著抑制bMECs 中该基因mRNA 的转录水平(p<0.05),而 8 μmol/L 的硒也能抑制该基因mRNA 的转录水平,但与模型组相比无统计学差异(p>0.05)(图1A)。
RIP2 mRNA 检测结果显示,在S.aureus感染6 h 后,与对照组相比,模型组细胞中RIP2 mRNA的转录水平显著上升(p<0.01)。与模型组相比,实验组中2 μmol/L 的硒能极显著抑制bMECs 中该基因 mRNA 的转录水平(p<0.01),而 4 μmol/L 和 8 μmol/L的硒能显著抑制该基因mRNA 的转录水平(p<0.05);在S.aureus感染8 h 后,与对照组相比,模型组细胞中RIP2 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,实验室 2 μmol/L、4 μmol/L 和8 μmol/L 的硒均能显著抑制bMECs 中该基因mRNA的转录水平(p<0.05);在S.aureus感染 10 h 后,与对照组相比,模型组细胞中RIP2 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L和 4 μmol/L 的硒均能显著抑制 bMECs 中该基因mRNA 的转录水平(p<0.05),而 8 μmol/L 的硒能抑制该基因mRNA 的转录水平,但与模型组相比无统计学差异(p>0.05)(图1B)。
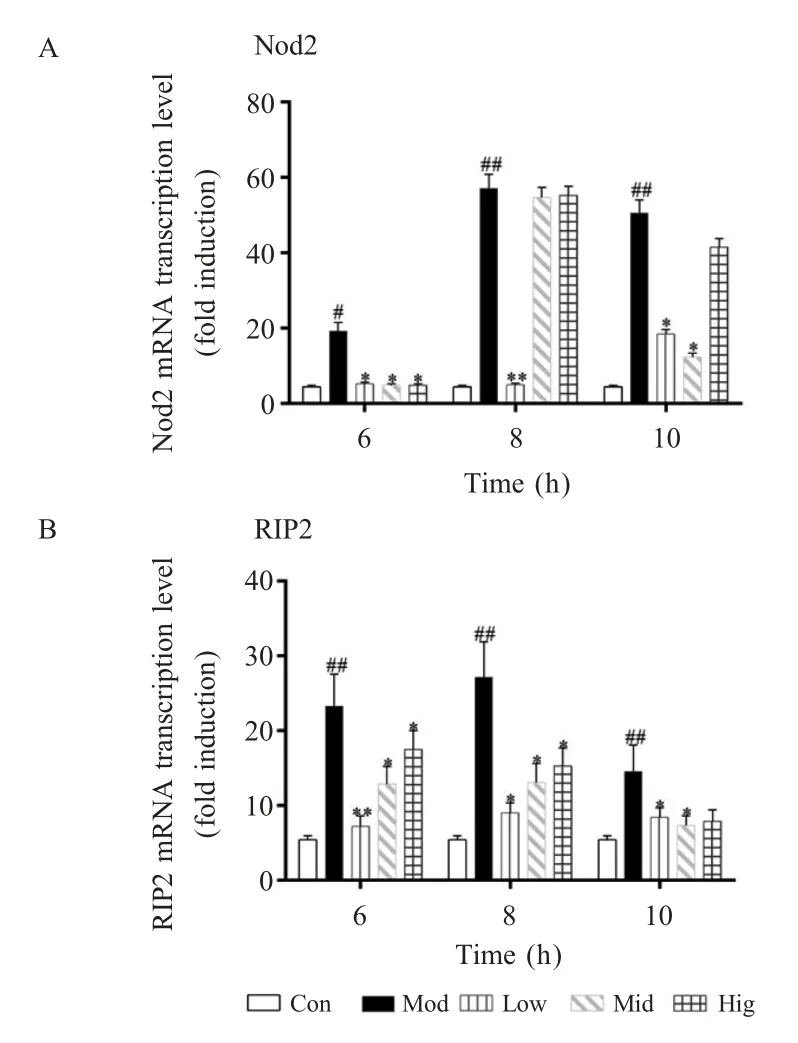
图1 硒对bMECs 中Nod2 和RIP2 mRNA 转录水平的影响Fig.1 Effects of Se on the transcriptional level of Nod2 and RIP2 mRNA
上述结果表明:在孵育时间较短的情况下(6 h)低浓度的硒(2 μmol/L)即可有效调控 Nod2 和RIP2 mRNA 的转录水平,但随着孵育时间的延长(8 h 和10 h)高浓度的硒效果开始减弱。2.2 硒对MAPK 信号通路关键因子JNK mRNA 转录水平的影响 本实验利用qPCR 方法对MAPK 信号通路关键因子JNK 的mRNA 转录水平进行了检测。结果显示,在S.aureus感染6 h 后,与对照组相比,模型组细胞中JNK mRNA 的转录水平显著上升(p<0.01)。2 μmol/L、4 μmol/L 和 8 μmol/L 的硒均能抑制bMECs 中该基因mRNA 的转录水平,但与模型组相比无统计学差异(p>0.05);在S.aureus感染8 h 后,与对照组相比,模型组细胞中JNK mRNA 的转录水平极显著上升(p<0.01),2 μmol/L 的硒能抑制bMECs 中该基因mRNA 的转录水平,但与模型组相比无统计学差异(p>0.05),而与模型组相比,4 μmol/L 和 8 μmol/L 的硒均能显著抑制 bMECs中该基因 mRNA 的转录水平(p<0.05);在S.aureus感染10 h 后,与对照组相比,模型组细胞中JNK mRNA 的转录水平极显著上升(p<0.01),与模型组相比,2 μmol/L、4 μmol/L 和 8 μmol/L 的硒均能显著抑制bMECs 中该基因mRNA 的转录水平(p<0.05)(图2)。结果表明:硒能抑制JNK mRNA 的转录水平,并且这种抑制作用存在时间依赖性,在孵育时间较短(6 h 和8 h)的情况下,高浓度的硒(8 μmol/L)才具备抑制相关基因表达的效果,而随着孵育时间的增加(10 h)低浓度的硒也可发挥该抑制作用。
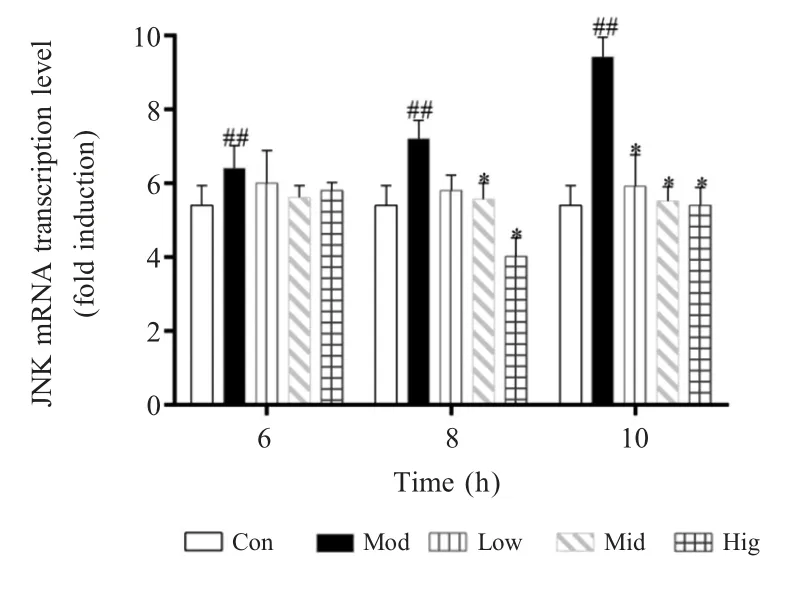
图2 硒对bMECs 中JNK mRNA 转录水平的影响Fig.2 Effects of Se on the transcriptional level of JNK mRNA
2.3 硒对 mTORs 信号通路关键因子 AKT 和mTOR mRNA 转录水平的影响本试验利用qPCR方法对mTORs 信号通路关键因子AKT 和mTOR 的mRNA 转录水平进行了检测。AKT mRNA 检测结果显示,在S.aureus感染6 h 后,与对照组相比,模型组细胞中AKT mRNA 的转录水平显著上升(p<0.01)。2 μmol/L 和 4 μmol/L 的硒均能抑制 bMECs 中该基因mRNA 的转录水平,但与模型组相比无统计学差异(p>0.05),与模型组相比,8 μmol/L 的硒能显著抑制 bMECs 中该基因 mRNA 的转录水平(p<0.05);在S.aureus感染8 h 后,与对照组相比,模型组细胞中AKT mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 和 4 μmol/L 的硒均能显著抑制 bMECs 中该基因 mRNA 的转录水平(p<0.05),而 8 μmol/L 的硒能极显著抑制 bMECs 中该基因mRNA 的转录水平(p<0.01);在S.aureus感染 10 h后,与对照组相比,模型组细胞中AKT mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 的硒能显著抑制 bMECs 中该基因mRNA的转录水平(p<0.05),而 4 μmol/L 和 8 μmol/L 的硒均能极显著抑制bMECs 中该基因mRNA 的转录水平(p<0.01)(图3A)。
mTOR mRNA 检测结果显示,在S.aureus感染6 h 后,与对照组相比,模型组细胞中mTOR mRNA的转录水平极显著上升(p<0.01)。2 μmol/L 和 4 μmol/L的硒均能抑制bMECs 中该基因mRNA 的转录水平,但与模型组相比无统计学差异,而与模型组相比,8 μmol/L 的硒能显著抑制bMECs 中该基因 mRNA的转录水平(p<0.05);在S.aureus感染8 h 后,与对照组相比,模型组细胞中mTOR mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 和4 μmol/L 的硒均能显著抑制bMECs 中该基因mRNA的转录水平(p<0.05),而 8 μmol/L 的硒能极显著抑制 bMECs 中该基因 mRNA 的转录水平(p<0.01);在S.aureus感染10 h 后,与对照组相比,模型组细胞中 mTOR mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 的硒能显著抑制bMECs 中该基因 mRNA 的转录水平(p<0.05),而 4 μmol/L 和8 μmol/L 的硒均能极显著抑制bMECs 中该基因mRNA 的转录水平(p<0.01)(图3B)。
上述结果表明:硒能抑制AKT 和mTOR mRNA的转录水平,并且抑制作用存在时间依赖性,在孵育时间较短(6 h)的情况下,高浓度的硒(8 μmol/L)才具备抑制相关基因转录的效果,而随着孵育时间的增加(8 h 和10 h)低浓度的硒也可发挥该抑制作用。
2.4 硒对炎症细胞因子IL-8 和IL-10 mRNA 转录水平的影响本实验通过qPCR 方法对炎症细胞因子IL-8 和IL-10 mRNA 转录水平进行了检测。IL-8 mRNA 检测结果显示,在S.aureus感染6 h 后,与对照组相比,模型组细胞中IL-8 mRNA 的转录水平显著上升(p<0.01)。与模型组相比,4 μmol/L 的硒能显著抑制bMECs 中该基因 mRNA 的转录水平(p<0.05),而 8 μmol/L 的硒能显著抑制 bMECs 中该基因 mRNA 的转录水平(p<0.01);在S.aureus感染8 h 后,与对照组相比,模型组细胞中IL-8 mRNA的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L、4 μmol/L 和 8 μmol/L 的硒能均能显著抑制 bMECs 中该基因 mRNA 的转录水平(p<0.05);在S.aureus感染10 h 后,与对照组相比,模型组细胞中IL-8 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L、4 μmol/L 和 8 μmol/L 的硒均能显著抑制bMECs 中该基因mRNA 的转录水平(p<0.05)(图 4A)。
IL-10 mRNA 检测结果显示,在S.aureus感染6 h后,与对照组相比,模型组细胞中IL-10 mRNA 的转录水平显著上升(p<0.01)。与模型组相比,2 μmol/L、4 μmol/L 和 8 μmol/L 的硒能均能显著抑制 bMECs中该基因 mRNA 的转录水平(p<0.05);在S.aureus感染8 h 后,与对照组相比,模型组细胞中IL-10 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L 的硒能显著抑制bMECs 中该基因mRNA 的转录水平(p<0.05),而 4 μmol/L 和 8 μmol/L的硒能极显著抑制bMECs 中该基因mRNA 的转录水平(p<0.01);在S.aureus感染 10 h 后,与对照组相比,模型组细胞中IL-10 mRNA 的转录水平极显著上升(p<0.01)。与模型组相比,2 μmol/L、4 μmol/L和8 μmol/L 的硒均能极显著抑制bMECs 中该基因mRNA 的转录水平(p<0.01)(图 4B)。
结果表明:硒能抑制炎症细胞因子IL-8 和IL-10 mRNA 的转录水平,并且抑制作用在孵育时间较短的情况下(6 h)即可表现出浓度的依赖性,随着孵育时间的增加(8 h 和10 h)各浓度的硒均可表现出极佳的抗炎效果。
3 讨 论
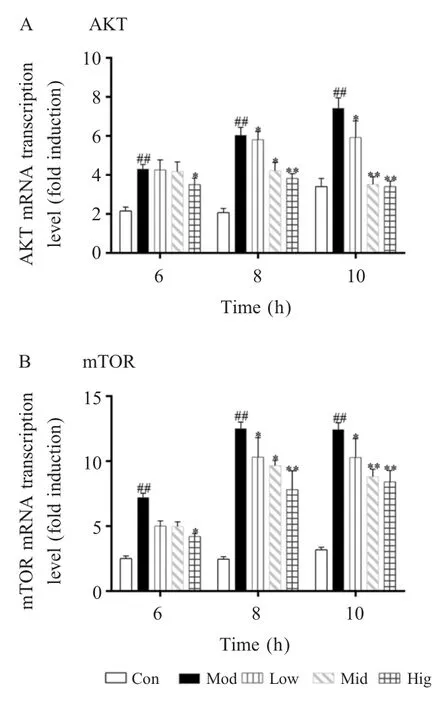
图3 硒对bEMCs 中AKT 和mTOR mRNA 转录水平的影响Fig.3 Effects of Se on the transcriptional level of AKT and mTOR mRNA

图4 硒对炎症细胞因子IL-8 和IL-10 mRNA 转录水平的影响Fig.4 Effects of Se on the transcriptional level of IL-8 and IL-10 mRNA
Nod2 和RIP2 是 Nod2 信号通路中的核心蛋白[8]。其中Nod2 蛋白是近年来发现的一种新的细胞内PRR,研究证明Nod2 蛋白可以识别多种类型的细菌肽聚糖[9]。活化的Nod2 蛋白主要通过CARD-CARD结构域之间的相互作用与RIP2 分子结合,并以RIP2 作为关键结合蛋白进一步诱导炎症反应[10]。适当的炎症对机体的免疫细胞有益,但严重的炎症会损害动物体的组织器官[11]。本研究发现bMECs 经S.aureus诱导 10 h 内,Nod2 信号通路中的 Nod2 和RIP2 基因被连续激活,表明Nod2 信号通路在S.aureus早期感染中起着重要的作用。此外,经过了6 h孵育后,低浓度的硒就表现出了抑制Nod2 和RIP2基因转录效果,这也间接反映出了模式识别受体Nod2 及其下游RIP2 在应对外界刺激过程中的灵敏性,随着孵育时间的延长高浓度的硒效果反而减弱,这可能与硒的毒性作用有关,在实验过程中即使8 μmol/L 的硒对细胞整体未产生毒性作用,但其对PRR 的毒性却是不可忽略的。
Nod2 信号通路主要可通过激活 MAPK 和mTOR 信号通路进一步诱导炎症反应。MAPK 信号通路是上皮细胞免疫系统的重要组成部分,而MAPK 蛋白是炎症调节过程中的关键因子[12]。MAPK 蛋白可分为 ERK、P38 和 JNK 3 个亚群[13]。研究发现硒对MAPK 蛋白中的p38 和ERK 的磷酸化有一定的影响[9]。但硒对MAPK 蛋白中JNK 影响的研究尚未见报道。本研究发现在孵育时间较短的情况下,只有高浓度的硒才能显著抑制JNK 基因mRNA 的转录水平,这可能与Nod2 信号通路与MAPK 信号通路的灵敏性相关,但随着孵育时间的延长低浓度的硒即可表现出抑制效果。
AKT 蛋白是连接MAPK 和mTOR 信号通路的核心蛋白[14]。活化的AKT 蛋白通过磷酸化下游的一些酶、激酶、转录因子等来调节细胞功能[15]。本研究采用qPCR 检测发现硒能抑制AKT mRNA 的转录水平,与对JNK 的调控效果相似,硒对AKT mRNA 的调控同样表现出时间效应关系。
mTOR 是AKT 的重要底物之一。AKT 可直接磷酸化mTOR 的SER 位点,激活mTOR 及其下游信号通路[16]。本研究采用qPCR 检测发现S.aureus诱导了mTOR 的转录,而硒能抑制mTOR 的转录。
IL-8 和IL-10 细胞因子具有广泛的生物学活性,可促进靶细胞的增殖分化,增强抗感染,促进或抑制其它细胞因子和膜表面分子的表达,促进炎症进程等[17]。本研究发现S.aureus显著增加了bMECs IL-8 和IL-10 mRNA 的转录水平,在孵育6 h 后硒能显著降低S.aureus诱导的 bMECs 中 IL-8 和 IL-10 mRNA 的转录水平,同时这种作用具有浓度效应关系,随着孵育时间的延长各浓度的硒均可显著抑制IL-8 和IL-10 mRNA 的转录从而达到抗炎效果。
总之,在奶牛乳腺炎过程中,S.aureus可通过诱导bMECs Nod2/MAPK/mTOR 等多条信号通路而触发炎症反应。但硒可通过抑制炎症相关信号通路中关键因子mRNA 的转录水平来减轻炎症对bMECs的损伤,限于PRR Nod2 及其下游RIP2 对硒毒性作用的敏感性,使用低浓度的硒(2 μmol/L)延长孵育时间效果最佳。
猜你喜欢
生殖医学杂志(2022年10期)2022-10-19
世界科学技术-中医药现代化(2022年2期)2022-05-25
中风与神经疾病杂志(2021年9期)2021-11-08
中老年保健(2021年5期)2021-08-24
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年4期)2021-07-31
感染、炎症、修复(2021年1期)2021-07-28
感染、炎症、修复(2021年1期)2021-07-28
中小学德育(2020年11期)2020-03-18
标记免疫分析与临床(2016年9期)2016-11-21
