
生鲜牛乳中金黄色葡萄球菌的鉴定及耐药分子特征研究
2019-11-22 03:08:00吴俐勤裘罕琦周沁怡戴贤君夏效东
中国预防兽医学报 2019年9期
张 玲,常 江,吴俐勤,裘罕琦,周沁怡,戴贤君,杨 华,夏效东,唐 标*
(1.西北农林科技大学食品科学与工程学院,陕西杨凌712100;2.浙江省农业科学院农产品质量标准研究所,浙江杭州310021;3.中国计量大学现代科技学院,浙江杭州310018;4.南京农业大学动物医学院,江苏南京210095)
金黄色葡萄球菌(Staphylococcus aureus)是一种常见的革兰氏阳性食源性致病菌。因肠毒素、溶血毒素、杀白细胞素等外毒素的产生,而经常引起食物中毒并在人类和动物中引起各种感染[1]。目前,微生物鉴定方法主要为生化鉴定和分子生物学鉴定。生化鉴定对细菌的培养状态有一定的要求,而分子生物学鉴定过程较繁琐,技术要求高。基质辅助激光解析电离飞行时间质谱(Matrix assisted laser desorption ionization time of flight mass spectrometry,MALDI-TOF MS)是一种基于蛋白指纹图谱进行微生物鉴定的新型技术[2-3],与生化鉴定和分子生物学鉴定相比,MALDI-TOF MS 鉴定不依赖细菌的生化反应,无需提取DNA,具有操作简单、通量高、灵敏度高、试剂耗材经济等优势,可以快速的区分和鉴定同一培养平板上未知细菌[4-5]。目前有研究表明MALDI-TOF MS 已经成功应用于沙门氏菌、大肠杆菌、阪崎克罗诺肠杆菌、S.aureus、弯曲杆菌等多种食源性致病菌的鉴定[6-7]。MALDI-TOF MS 作为一种新型的分类方法,具有快速、成本低等优点,在细菌的聚类分析上同样具有很大的潜力[8]。菌株的获得是细菌耐药性研究的前提,质谱法对于纯培养物的特定病原菌的鉴定和未知病原菌的发现具有明显优势,是动物源致病菌菌株库建设的重要技术支撑。
可移动元件是细菌介导耐药基因转移的重要载体,包括转座子、质粒和噬菌体等[9]。质粒可以介导多个耐药基因对不同种类的抗生素耐药,在S.aureus的耐药性传播中起主要作用。在动物养殖和治疗过程中,过量、不合理的使用抗生素,对耐药基因具有选择压力,从而增加了耐药质粒转移和重组概率,进而诱发S.aureus多重耐药的产生。特别是耐甲氧西林S.aureus(MRSA),作为一种重要人畜共患病原体,不仅增加治疗成本,同时威胁公共卫生安全,是全球范围的主要健康威胁之一[10-11]。因此,对养殖场的S.aureus的兽药敏感性监测非常必要,是指导养殖场合理用药的重要依据。
本研究从养殖场生鲜乳中利用MALDI-TOF MS快速分离鉴定了S.aureus,并对其进行了最低抑菌浓度(MIC)测定。从全基因组的角度,分析了获得性耐药基因,并对携带质粒进行了比对分析,初步解释了介导其耐药性的分子基础,为奶牛场合理用药和控制抗药性提供重要参考。
1 材料与方法
1.1 主要实验材料浙江省杭州市某奶牛场99 份生鲜牛乳样品(50 份未患病奶牛牛乳、49 份患有隐腺乳房炎奶牛的牛乳);质控菌株为ATCC 29213(本实验室保藏);缓冲蛋白胨水(BPW)、7.5 % NaCl肉汤、Baird-Parker 琼脂、LB 琼脂、卵黄亚碲酸钾增菌剂购自北京陆桥技术有限公司;MALDI-TOF MS 基 质 : α- 氰 基 -4 羟基肉桂酸 (4-Hydroxy-α-cyanocinnamic acid,HCCA)购自 Sigma 公司;基质溶剂[乙腈(Acetonitrile,ACN):水:三氟乙酸(Trifluoroacetic acid,TFA)的比例为 50:47.5:2.5]、蛋白标准品BTS (Bacteria Testing Standards)购自德国布鲁克公司;2×TaqMaster Mix 和 DNA Marker 购自TaKaRa 公司;细菌基因组DNA 提取试剂购自上海捷瑞生物工程有限公司;Gel-red 核酸染料、50×TAE 缓冲液、葡萄球菌溶菌酶(Lysostaphin)购自生工生物工程(上海)股份有限公司;96 孔肉汤微量稀释法药敏板购自上海星佰生物技术有限公司;基质辅助激光解吸电离飞行时间质谱仪购自德国布鲁克·道尔顿公司。
基因检索号:S.aureusSABHZ053 和SABHZ079基因组序列的检索号分别为QZVV00000000、QZVW00000000;质粒pSA053-1、pSA053-2、pSABHZ079-1、pSABHZ079-2 和 pSABHZ079-3 的检索号分别为 NZ_CM011642、NZ_CM011643、NZ_CM011644、NZ_CM011645 和 NZ_CM011646。
1.2 S.aureus 的分离参考GB 4789.10-2016《食品安全国家标准食品微生物学检验金黄色葡萄球菌检验》中的方法分离S.aureus。取1 mL 生鲜牛乳样品加入10 mL 7.5 %的NaCl 肉汤中,37 ℃培养24 h;将增菌液划线于Baird-Parker 琼脂(每 95 mL加5 mL 的卵黄亚碲酸钾增菌剂),37 ℃培养24 h;挑取带有透明圈和沉淀环的黑色单菌落-划线于LB琼脂上37 ℃培养24 h,其他疑似菌落于相同条件下划线培养。
1.3 菌落的MALDI-TOF MS 鉴定分析将可疑单菌落均匀涂布至MALDI 靶板上的小孔中,加入1 μL 70 %的甲酸溶液破壁,自然晾干,再加入1 μL HCCA 基质溶液覆盖,自然晾干、结晶。使用仪器前,先用标准品BTS 校正,再进行待测菌图谱的采集。利用MALDI Biotyper RTC 软件[12]进行样品指纹图谱的自动采集及分析,将采集的指纹图谱与数据库中的细菌标准图谱进行比较,并给出相应的分值,分值越高鉴定准确的可信度越高,分值为2.3~3.0 表示完全可靠的鉴定到种水平,分值为2.0~2.299 表示鉴定到种水平,分值为1.7~1.9 表示鉴定到属水平,分值在1.7 以下表示没有可信的鉴定结果。根据Biotyper RTC 获得的高质量图谱,利用MALDI Biotyper FlexAnalysis 软件先对其指纹图谱进行基线校正、平滑,然后通过MALDI BioTyper 主成分分析(PCA)树状图构建方法,采用默认参数构建聚类分析树。
1.4 S.aureus 16S rDNA 测序分析参照细菌基因组 DNA 提取试剂盒说明书提取S.aureus基因组DNA,利用溶葡菌酶破壁。抽取的基因组DNA 通过琼脂糖凝胶电泳和Nanodrop ONE 进行检测,质量合格后于-20 ℃保存备用。利用16S rDNA 基因通用引物(27F:AGAGTTTGATCMTGGCTCAG/1525R:AAGGAGGTGWTCCARCC)对 1.3 鉴定的S.aureus进行 PCR 扩增。PCR 反应条件为 94 ℃ 5 min;98 ℃10 s、57 ℃ 20 s、72 ℃ 60 s,共 35 个循环;PCR产物经琼脂糖凝胶电泳检测后,由杭州擎科梓熙生物技术有限公司测序,测序结果通过BLAST 比对鉴定。将测得的16S rDNA 基因序列用Clustal W 软件进行对位分析,采用MEGA X 软件以最大似然法进行系统发育树的构建,进化树分支模式的稳定性分析采用bootstrap 方法,重复次数为1 000 次。
1.5 S.aureus 的药敏试验根据美国临床实验室标准化委员会(Clinical And Laboratory Standards Institute,CLSI)2015 版 M100-S25 文件[13],采用肉汤微量稀释法测定S.aureus对18 种抗菌药物的MIC,质控菌株为ATCC 29213。同时利用mecA基因引物(mecAF: TCATAGCGTCATTATTCCAGG/mecAR:AATTTGTCTGCCAGTTTCTCCT)对 5 株S.aureus进行PCR 扩增鉴定,对两种检测结果进行对比分析。1.6 两株S.aureus的全基因组测序及分析 利用Illumina Hiseq 测序平台对两株耐甲氧西林的S.aureus(SABHZ053 和SABHZ079)进行全基因组测序,利用Velvet 软件对序列组装。利用ResFinder 3.1 软件(https://cge.cbs.dtu.dk/services/ResFinder/)对耐药基因进行预测,利用PlasmidFinder 2.0 软件(https://cge.cbs.dtu.dk/services/PlasmidFinder/)进行质粒复制子类型的预测。预测到的质粒序列经PCR、测序和拼接后获得完整质粒序列,利用EasyFig 和BRIG 软件对质粒序列比较分析。
2 结 果
2.1 S.aureus 的分离结果从99 份生鲜牛乳样品中共分离得到5 株S.aureus(分离率5.1 %),均分离自患有隐性乳房炎的奶牛,未患病奶牛未分离到S.aureus。表明奶牛生鲜乳中S.aureus污染风险低,奶牛乳房炎不仅仅是由S.aureus引起的。
2.2 不同形态葡萄球菌的MALDI-TOF MS 鉴定利用MALDI-TOF MS 对Baird-Parker 琼脂上的细菌进行鉴定,与S.aureus具有相近形态的代表性菌落得到快速区分:SABHZ053 为S.aureus,其菌落为圆形、黑色、有光泽,周围有混浊带,外层有透明圈;SABHZ055 为表皮葡萄球菌(S.epidermidis);SABHZ092 为克氏葡萄球菌(S.kloosii);SABHZ088为马胃葡萄球菌(S.equorum);SABHZ041 为产色葡萄球菌(S.chromogenes);SABHZ046 为粪肠球菌(Enterococcus faecalis);SABHZ001 为模仿葡萄球菌(S.simulans);SABHZ026 为溶血性葡萄球菌(S.haemolyticus)(图1)。

图1 Baird-Parker 琼脂上不同形态的菌落Fig.1 Different forms of colonies on Baird-Parker agar
2.3 16S rDNA 与 MALDI BioTyper PCA 系统发育树比较将分离获得的5 株S.aureus、7 株其他葡萄球菌进行 16S rDNA 聚类分析(图2A),同时采用MALDI-TOF MS 聚类分析(图2B),粪肠球菌作为外类群。两种方法获得的树形具有较高的一致性,其中S.aureus均聚成一个分支,并且聚类分析树在质谱鉴定后即可获得,速度上具有绝对优势,显示出了MALDI-TOF MS 聚类分析的潜力。
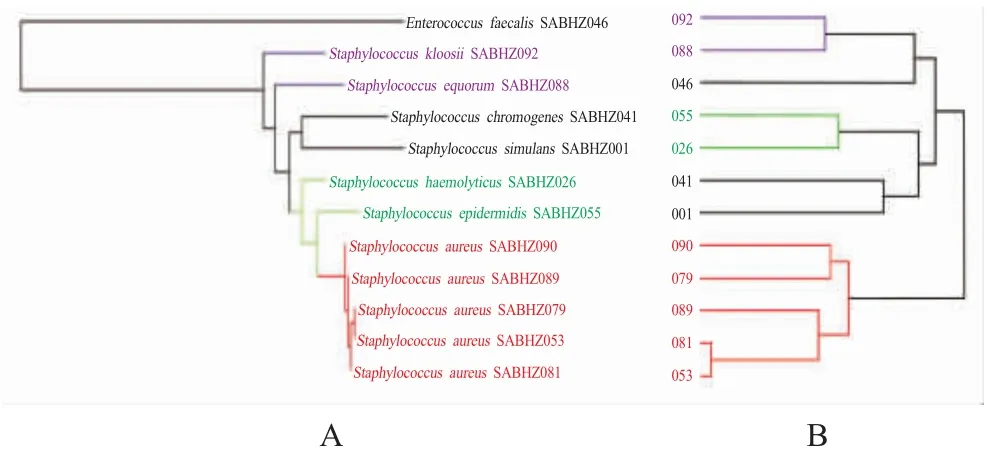
图2 16S rDNA 最大似然法系统发育树(A)和质谱PCA 树状图(B)比较Fig.2 16S rDNA maximum likelihood tree (A)and MALDI BioTyper PCA dendrogram (B)
2.4 S.aureus 的药敏试验PCR 扩增证实 SABHZ079、SABHZ053 两株菌mecA基因阳性,均为耐甲氧西林S.aureus(MRSA),其中SABHZ079 表现出7 重耐药,SABHZ053 对2 种抗生素耐药;但5 株S.aureus均对青霉素(PEN)耐药(表 1)。
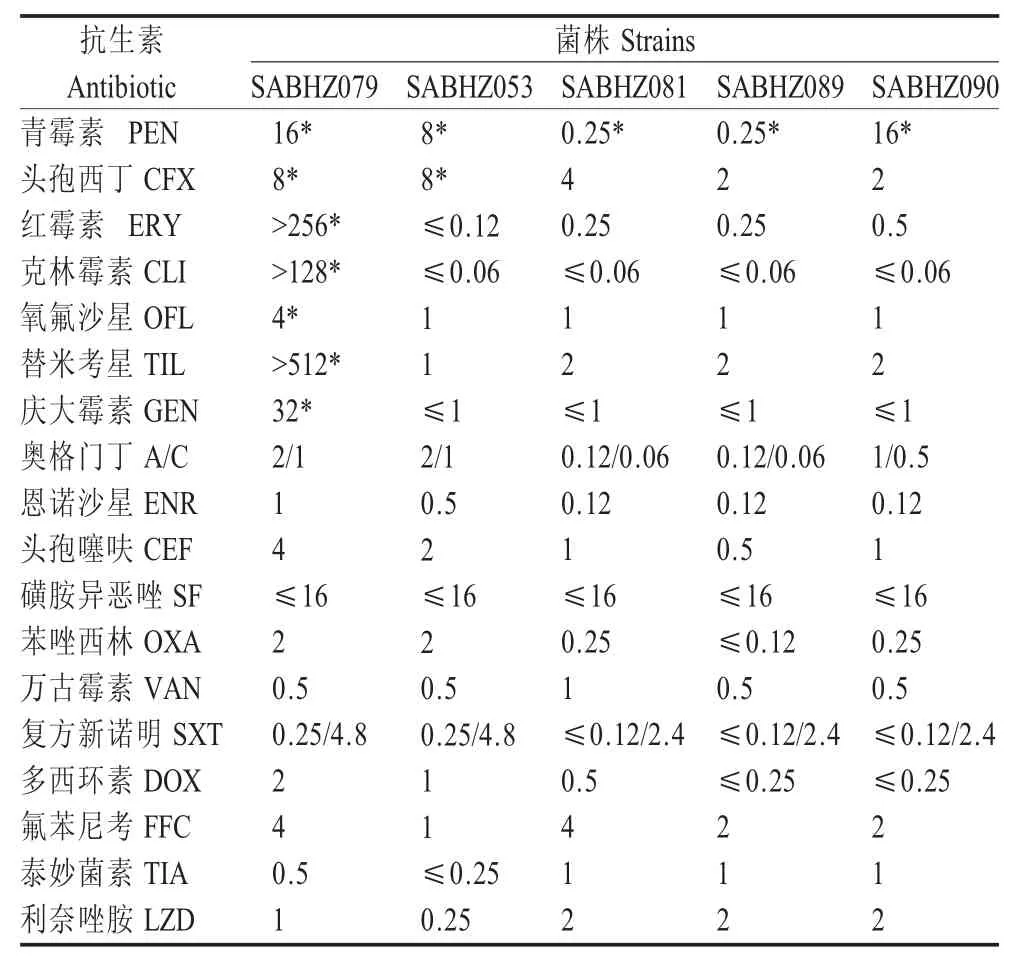
表 1 5 株 S.aureus 的 MICTable 1 MICs of five Staphylococcus aureus strains
2.5 菌株SABHZ053 和SABH079 的质粒序列比较分析对菌株SABHZ053 和SABHZ079 进行全基因组测序,测序深度分别为967X、887X。组装获得的基因组通过预测,两株菌的MLST 基因型均为ST965。前者有2 个质粒,后者有3 个质粒(图3)。
通过ResFinder 预测两株分离株的耐药基因分别为aph(3')-III、ant(6)-Ia、blaZ、mecA、tet(K)和blaZ,aph (3')-III、ant (6)-Ia、aph (2'')-Ia、mecA、blaZ、erm(B)、erm(C)、tet(K)。SABHZ053 的两个耐药基因tet(K)、blaZ分别存在于质粒 pSA053-1(NZ_CM011642,4 439 bp)、pSA053-2(NZ_ CM011643,34 609 bp)中,而 SABHZ079 的 5 个耐药基因aph(2'')-Ia、blaZ、erm(B)、erm(C)和tet(K)分别位于质粒 pSABHZ079-1 (NZ_CM011644, 39 227 bp)、pSABHZ079-2 (NZ_CM011645, 2 473 bp)、 pSABHZ079-3(NZ_CM011646,4 439 bp)中(表 2)。存在于该两株菌的质粒pSABHZ079-3 和pSA053-1 中的tet(K)基因介导四环素类耐药、pSABHZ079-1 和pSA053-2 中的blaZ基因对青霉素耐药[14],区别在于菌株 SABH079 的质粒 pSABHZ079-1 中的aph(2'')-Ia基因对庆大霉素耐药[15],质粒pSABHZ079-1 中的erm(B)基因和质粒pSABHZ079-2 中的erm(C)基因对大环内酯类ERY、CLI、TIL 耐药起到主要作用[16-17](表2)。结果显示,aph(2'')-Ia与erm(B)基因在质粒pSABHZ079-1 中相对质粒pSA053-2 是插入,在该两个基因前可以预测到转座酶,作为一个转座单元介导大环内脂和氨基糖苷类药物的耐药(图3)。
通过与GenBank 数据库比对后发现,质粒pSABHZ079-1 和pSA053-2 在已登录的核酸序列中不存在与该两个质粒完全匹配的质粒。将检索获得的同源质粒与pSABHZ079-1 进行比对分析,发现该质粒可明显分为骨架I 和骨架II 两部分(图4)。质粒pSA053-2、pN315(AP003139,24 653 bp)、pWBG752(GQ900394, 24 654 bp)、 pNCCP14558 (CP013954,24 729 bp)、 pNCCP14562 (CP013956, 24 657 bp)、pTZ2162 (AB304512,35 380 bp)与 pSABHZ079-1 骨架I 的核酸序列同源性平均为99.8%(大小为24.6 kb),且均来源于S.aureus;质粒 pFORC59 (CP020355,35 269 bp)、 pFORC_039 (CP015818, 35 415 bp)、pNTUH_3874 (LC102479, 14 566 bp)、 pMCCL2(AP009486,80 545 bp)与该质粒骨架II 的核酸序列同源性平均为99.9 % (大小为14.5 kb),该骨架包含Tn551-erm(B)-aph(2'')-Ia转座单元。进一步研究发现质 粒 pN315、 pWBG752、 pNCCP14558、 pNCCP14562 与质粒的pSABHZ079-1 骨架 I 全长匹配;pNTUH_3874 与该质粒的骨架II 全长匹配,推断质粒pSABHZ079-1 可能是分离自临床的pNTUH_3874与pN315 或其相似质粒杂合而来(图4)。
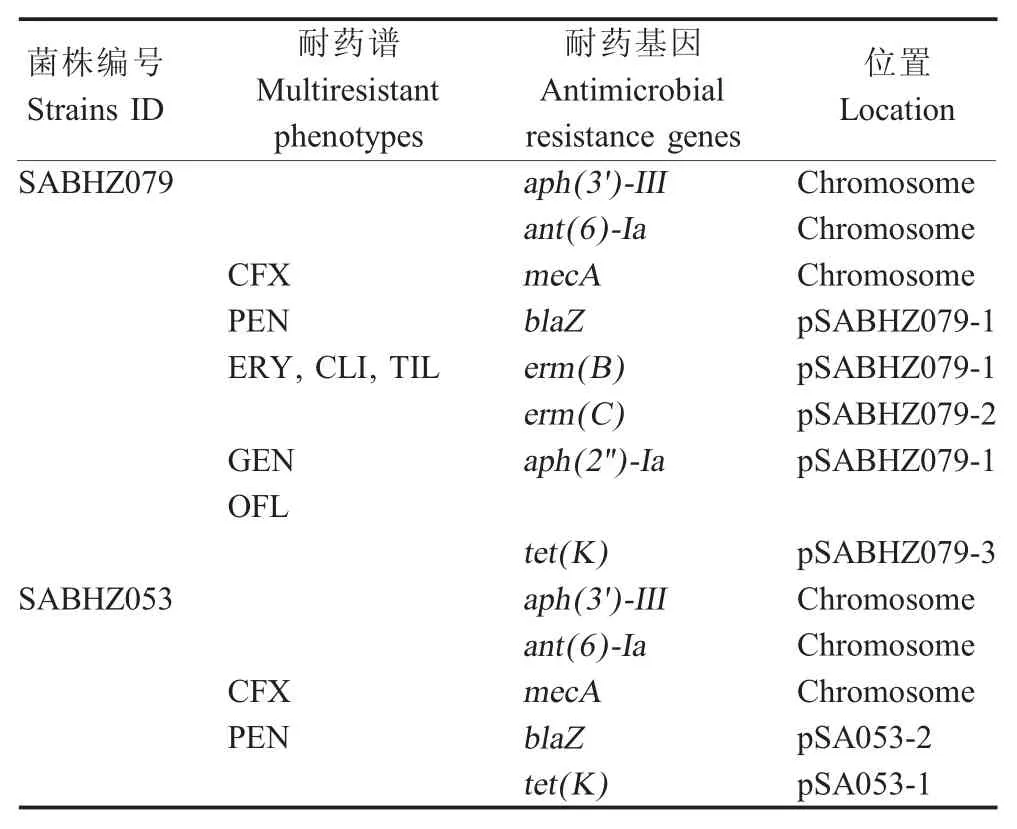
表2 菌株SABHZ053 和SABHZ079 的耐药谱与耐药基因Table 2 Antimicrobial resistance profiles and genes of strain SABHZ053 and SABHZ079

图3 菌株SABHZ053 和SABHZ079 耐药质粒的比较分析Fig.3 Comparative analysis of plasmids between SABHZ053 and SABHZ079 strains
3 讨 论
本研究从杭州某奶牛场生鲜乳中分离葡萄球菌,通过MALDI-TOF MS 快速鉴定得到5 株S.aureus,且均来自患有隐形乳房炎的奶牛。这表明该奶牛场S.aureus的污染程度不高,S.aureus的存在是导致奶牛患隐形乳房炎的原因之一。对其中5 株S.aureus和7 株葡萄球菌通过MALDI BioTyper 软件的PCA 分析进行了树形图的构建,发现该树形图和16S rDNA 系统发育树一致性较高。该方法在动物源病原菌的分离鉴定和分型过程中显示出速度优势。通过质谱鉴定,可以快速判断是否为目标菌株,同时也获得了其他非目标菌株的种属,有利于致病菌菌株库的构建。

图4 质粒pSABHZ079-1 与同源质粒的比较分析Fig.4 Comparative analysis of plasmid pSABHZ079-1 with homologous plasmids
药敏结果显示,5 株S.aureus均对青霉素耐药,其中 2 株S.aureus对头孢西丁耐药,其中 SABHZ079 对7 种抗生素耐药。养殖场抗生素的不合理使用容易引发致病菌多重耐药,不仅给动物治疗带来了困难,同时还会威胁人类健康。指导养殖场合理的使用抗生素是遏制细菌耐药性的关键一环,因此针对致病菌的药敏监测非常重要。
头孢西丁可以作为苯唑西林耐药性检测的替代物,可以根据头孢西丁结果判断细菌对苯唑西林敏感或耐药[13]。在此研究中分离获得的S.aureus通过药敏试验后,仅对头孢西丁具有抗性的菌株通过PCR 验证,确认两株为MRSA。分离得到的两株MRSA 菌株的基因型同为ST965,在临床样品中也发现存在这一类型[9]。两株菌含有一个相同的质粒,还存在与该两个质粒同源性较高的质粒,区别在于这两个质粒一个质粒含有耐药基因erm(B),一个含有aph(2'')-Ia的转座元件。相比菌株 SABHZ053,SABHZ079 多了一个2.4 kb 的质粒,携带有erm(C)。erm(B)和erm(C)均可以介导S.aureus对大环内脂类耐药[16-18],在本研究中两株菌对红霉素、克林霉素和替米考星耐药,上述两个基因哪一个占主导作用还不明确。另外,相比菌株SABHZ053,SABHZ079 插入的aph(2'')-Ia基因应该明确为介导庆大霉素耐药的因素。在质粒的同源性比较中发现,分离测序获得的pSABHZ079-1 质粒是全新的质粒,从质粒的比对来看,它由两个分离自人源样本的S.aureus全长质粒杂合而来。这进一步体现了该奶牛场S.aureus携带耐药质粒传播与变异的复杂性。表明质粒是造成该奶牛场S.aureus耐药的重要原因,而转座插入又增加了耐药基因传播的风险。
该实验过程中分离获得的其它葡萄球菌目前未做药敏实验,耐药情况未知,但其无疑是重要的耐药质粒或基因携带菌[19],与S.aureus会彼此水平传播耐药基因。本实验室将会针对奶牛场葡萄球菌的耐药组和质粒组进行深入研究。
猜你喜欢
今日农业(2021年11期)2021-08-13 08:53:24
兽医导刊(2019年1期)2019-02-21 01:14:06
食品科学(2018年10期)2018-05-23 01:27:28
兽医导刊(2015年9期)2016-01-04 12:00:06
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
云南畜牧兽医(2014年2期)2014-02-28 21:25:17
遗传(2014年3期)2014-02-28 20:58:49
世界科学(2014年8期)2014-02-28 14:58:31
