
2016年~2018年山东地区25株H9N2亚型禽流感病毒分子分析
2019-11-22 03:08陈照坤黄小莹马鑫鑫朱凤珠黄庆华古曙光杨少华许传田
中国预防兽医学报 2019年9期
陈照坤,鲁 梅,黄小莹,马鑫鑫,朱凤珠,黄庆华,古曙光,杨少华,许传田*,崔 宁*
(1.山东师范大学生命科学学院,山东济南250014;2.山东省农业科学院畜牧兽医研究所/山东省畜禽疫病防治与繁育重点实验室,山东济南250100;3.山东潍坊工程职业学院,山东青州262500;4.山东泰安肥城市畜牧兽医局;山东肥城271600;5.日照市东港区西湖镇畜牧兽医站,山东日照276815)
1994年在我国广东地区首次分离到H9N2 型禽流感病毒(AIV),随后在我国大部分地区家禽群体中广泛流行[1]。禽流感(AI)是由A 型流感病毒引起的一种禽类病毒性传染病。AIV 为单链负RNA 病毒,基因组由8 条单股负链RNA 构成。根据病毒表面糖蛋白(HA)和神经氨酸酶(NA)的抗原性差异,AIV可划分为18 个 HA 亚型和 11 个 NA 亚型[2-3];依据AIV 对SPF 鸡致病性的强弱,将其分为高致病性AIV(HPAIV)和低致病性 AIV(LPAIV)两种类型[4]。H9N2 AIV 属于LPAIV,但其广泛存在变异重组现象,且可以为H5N1、H7N9 及H10N8 等能够直接感染人并致死的AIV 提供部分甚至整套内部基因,潜在危害备受关注[5-7]。H9N2 AIV 可以作为“载体”将不同亚型的流感病毒从禽类传播给人类,对人类健康、养殖产业具有极大的威胁[8],被国际卫生组织(WHO)认为是潜在的、可能引起下一轮暴发的AIV。基于以上原因,本研究对分离到的25 株H9N2 AIV 的基因组序列和遗传进化关系进行了系统性分析,从分子水平上了解山东地区H9N2 AIV变异情况和导致其表现不同生物学特性的原因,为H9N2 AIV 跨物种传播感染哺乳动物提供分子预警材料。
1 材料与方法
1.1 主要实验材料疑似H9N2 AIV 的489 份病料样品采自山东及周边养殖场;9 日龄~11 日龄SPF鸡胚购自山东省农业科学院SPF 鸡场;DL2000 Marker、一步法RT-PCR 试剂盒、RNAiso 试剂、反转录酶、dNTP、rTaq酶、pMD18-T 载体试剂盒、DH5α 感受态细胞均购自TaKaRa 公司;胶回收试剂盒购自天根生化科技(北京)有限公司;青霉素(80 IU)、链霉素(100 IU)、1.2 %琼脂糖凝胶、无菌PBS、LB培养液、LB 固体培养基、SOC 溶液均由本实验室配制。
1.2 病毒分离与鉴定将疑似H9N2 AIV 的病料匀浆处理后离心取上清,接种9 日龄~11 日龄SPF 鸡胚,37 ℃孵化72 h,无菌收集对鸡红细胞有血凝性的尿囊液,提取病毒RNA,参考GenBank 中已登录的H9N2 AIV 序列,设计鉴定引物(表1),由上海生工生物工程技术服务有限公司合成。通过鉴别诊断引物鉴定H9N2 AIV,鉴定正确的H9N2 AIV 通过有限稀释法纯化3 代,纯化的病毒通过SPF 鸡胚扩增后,收集尿囊液分装作为种毒,-80 ℃保存。

表1 H9N2 AIV 鉴别检测及测序引物Table 1 Different identification and sequencing primers for H9N2 AIV
1.3 病毒各基因节段的PCR 扩增及克隆测序按照 RNAiso 按照试剂盒说明提取1.2 得到的 H9N2 AIV RNA 并反转录为cDNA。根据各个基因节段参考序列设计合成12 对H9N2 AIV 全基因组扩增引物(表1)进行扩增,经1.2 %琼脂凝胶电泳检测,胶回收,分别连接至pMD18-T 载体,转化,经菌液PCR鉴定阳性菌液送由上海生工生物工程技术服务有限公司测序。
1.4 序列分析利用DNAStar 软件将分段测序的基因节段拼接为完整的开放阅读框(ORF),利用MegAlign 软件对各节段重要位点的氨基酸进行比对分析,按照核苷酸同源性95 %以上作为一个分支,利用MAGE 4.0 软件制作8 个基因节段的核苷酸遗传进化树,分析分离株H9N2 AIV 的分子进化关系。
2 结果与讨论
2.1 病毒的分离与鉴定通过RT-PCR 从489 份疑似AI 病料样品中鉴定得到76 株H9N2 AIV,分离纯化获得25 株H9N2 AIV (表2),其中5 株来源于鸭,20 株来源于鸡,病料均来自山东周边的规模化养殖场的商品鸭、商品鸡,发病日龄在25 日龄~35日龄,商品鸭未免疫H9 疫苗,商品鸡7 日龄免疫H9 灭活苗。
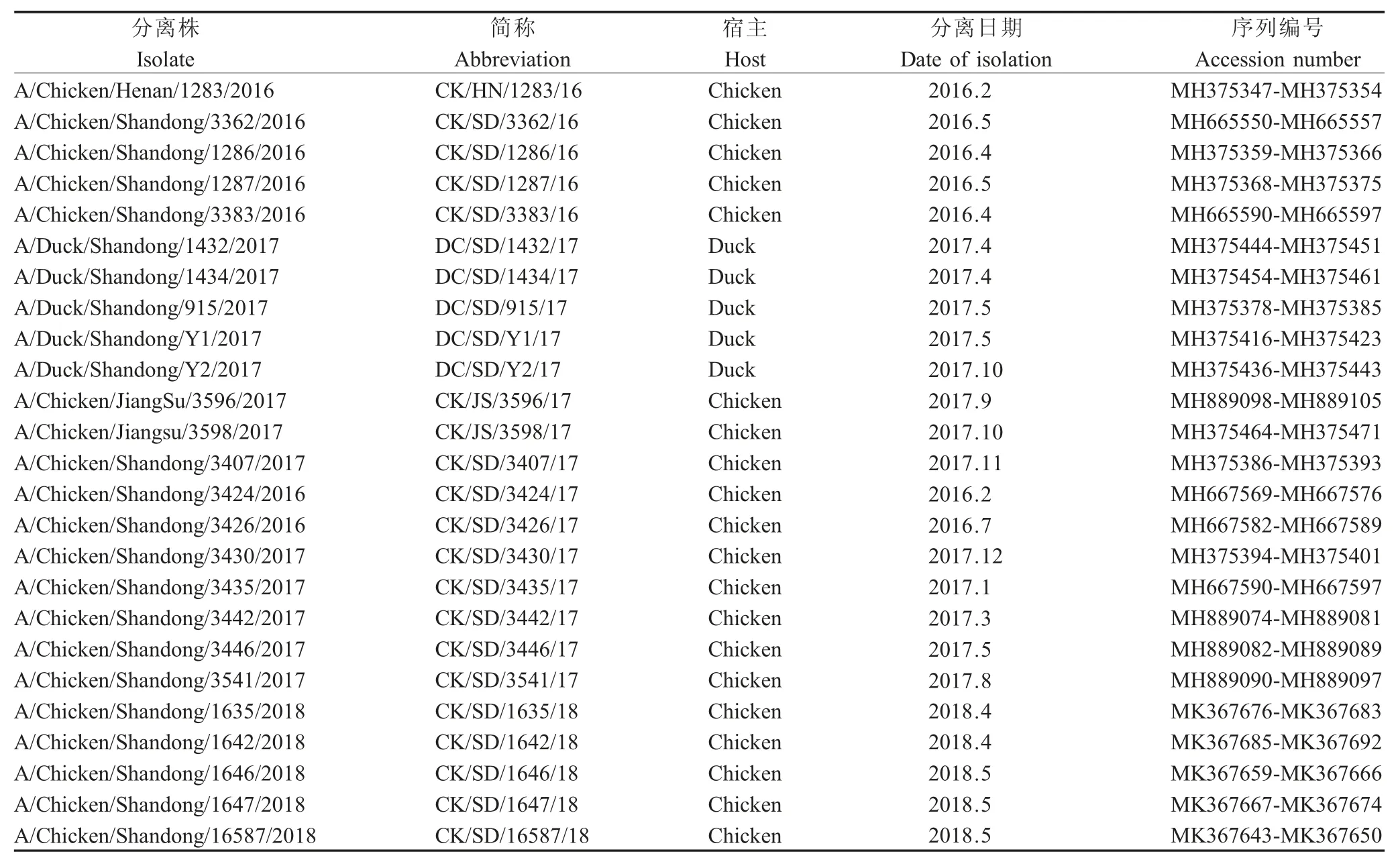
表2 25 株H9N2 AIV 分离背景信息Table 2 Isolation background information of 25 strains of H9N2 AIV
2.2 8 个基因节段遗传进化和关键氨基酸位点分析
2.2.1 HA 基因及关键位点氨基酸分析 25 株分离株HA 基因之间的同源性为92.8 %~99.9 %,与Y280-like 的同源性最高,达89.0 %~91.7 %。与已报道的2010年~2014年山东分离株同源性较高,说明近几年的山东地区H9N2 AIV 在核苷酸水平上变异不明显[9]。HA 基因系统进化树显示,25 株H9N2 AIV 均属于欧亚谱系,与Y280 分布在一个大分支,属于Y280-like(图1)。HA 基因裂解位点除了D/SD/Y1/17 是333PSRSNR↓GLF341外,其它 24 株均是333PSRSSR↓GLF341,表明 25 株H9N2 AIV 分离株HA 基因裂解位点仍符合LPAIV 的序列特征。25 株分离株的受体结合位点(RBS)部分关键氨基酸位点发生变异,共有6 株发生K149T、K149A 变异,20 株发生A198V、A198T、A198K 变异,25 株分离株均发生Q234L 变异,这种突变有易感染哺乳动物的趋势[10]。糖基化位点数目与位置形成新的变异趋势,25 株分离 株 在 29 ~31、 82 ~84、 141 ~143、 218 ~220、298~300、305~307、313~315、492~494、551~553 位出现潜在糖基化位点,21 株在218~300 位糖基化位点缺失,25 株分离株在313~315 位均出现潜在糖基化位点,这与分离自华南地区的H9N2 AIV 报道一致[11],与2012年上海发病鸡群中分离的3 株H9N2 AIV 变异类似[12]。以上结果说明我国的H9N2 AIV 病毒株变异趋势基本类似,这是否与疫苗的普遍应用引起的免疫选择压力有关,需要进一步证实,这些糖基化位点的变异对病毒致病性的影响也有待深入研究。
2.2.2 NA 基因及关键位点氨基酸分析 研究表明NA 蛋白在病毒的成熟、释放及感染新细胞方面均起着关键作用[13-14],因此对H9N2 AIV 的NA 基因变异分析的研究有着重要的意义。25 株H9N2 AIV HA 基因之间的同源性可达91.0 %~99.9 %,与Y280-like 核苷酸同源性相对较高,为90.3%~92.8%。NA 基因系统进化树显示,25 株H9N2 AIV 的NA基因均属于欧亚系,属于Y280-like 谱系(图1)。NA蛋白总共发现10 个潜在的糖基化位点,在69、86、146、200、234 位比较保守,在 264、368 和 402 位发生突变,60 位糖基化位点全部缺失。王守春等认为山东地区H9N2 AIV 的NA 蛋白具有失去402 糖基化位点、获得264 糖基化位点的趋势[15],本研究中的25 株分离株的NA 蛋白在402 位仅有3 个病毒株有糖基化位点,而在264 位共有11 个病毒株出现了糖基化位点,进一步证实了这一观点。这些糖基化位点的变化是否对病毒生物学特性造成影响有待进一步研究。 4 株分离株(DK/SD/Y1/17、DC/SD/Y2/17、DC/SD/1432/17、DC/S/1434/17)NA 蛋白的366、399 位红细胞结合位点未发生任何变异,其它位点处均发生不同程度的变化,366 ~373、399~404 位点氨基酸变异较大,431~433 位点处的氨基酸区域高度保守,25 株H9N2 AIV NA 蛋白均在63~65 位点缺失TEI。结果表明25 株H9N2 AIV NA 蛋白分离株NA 基因红细胞结合位点也发生不同程度的变异。
2.2.3 PB2、PB1、PA、NP、M、NS 基因的遗传进化分析 系统进化树显示,PB2 基因来自G9-like;PB1、PA、NP、NS 基因来自 F98-like;M 基因来自G1-like,6 个基因节段均属于欧亚谱系(图1),表明这几个基因节段是由不同进化方式而来。这说明近3年山东周边流行的H9N2 AIV 是由不同进化方式而来,这可能是近几年H9N2 AIV 在兽医临床案例中对家禽致病性和危害程度表现多样化的原因之一,引起这种差别的具体机制有待于进一步研究。
2.2.4 PB2、PB1、PA、NP、M、NS 关键位点氨基酸变异分析 研究表明,A 型流感病毒PB2 蛋白的E627、D701和S7143 个位点中任一位点发生突变均会导致病毒对小鼠的致病力增强[16-17]。本研究中25 株分离株中有 5 株(CK/SD/3424/16、CK/JS/3596/17、CK/SD/3442/17、 CK/SD/3541/17、 CK/SD/16587/18)PB2 氨基酸发生了E627V 的变异,1 株(CK/JS/3598/17)发生了S714C 的变异,表明山东地区的H9N2 AIV开始发生变异;PA 蛋白中672L 对病毒气溶胶传播特性有一定影响[16-17],25 株分离株PA 蛋白第672 位氨基酸均为L,表明这些分离株均具有通过气溶胶传播的能力;M2 蛋白中发生S31N 变异会使流感病毒对金刚烷胺类药物的抵抗力增强[16-17],25 株分离株M2 蛋白序列中均发生S31N 的变异,表明这些分离株均有对金刚烷胺类药物的抵抗力增强的可能性;NS1 蛋白第D92、E97和A149位氨基酸均未发生变异,干扰素可能仍然对这25 株分离株起作用。以上结果表明,近几年的H9N2 AIV 分离株对哺乳动物的致病力、横向传播以及耐药性可能有增强的趋势。
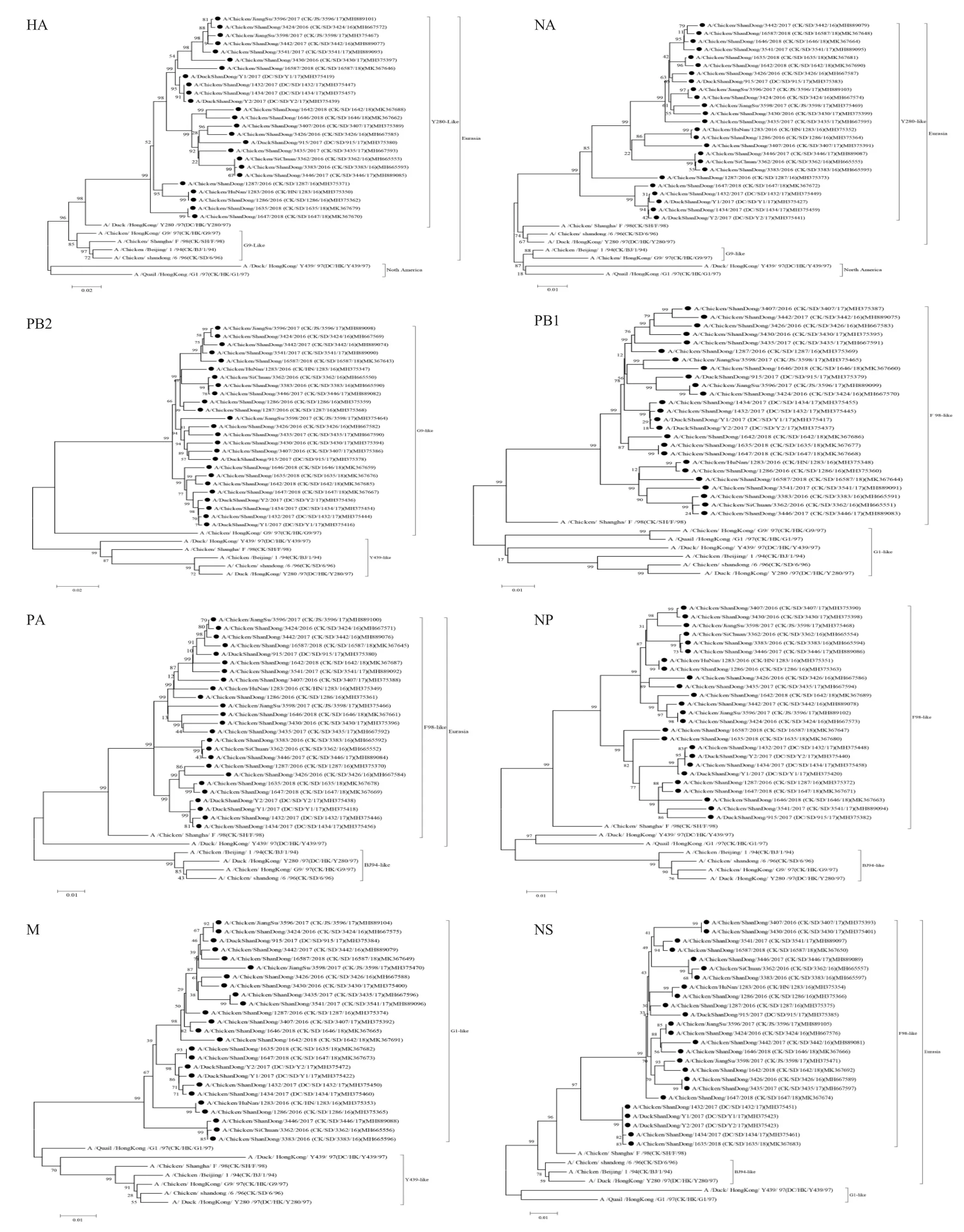
图1 25 株分离株的8 个基因节段与参考序列构建的遗传进化树Fig.1 Genetic evolution tree based on 8 gene fragments of 25 isolates and reference sequences
总之,近3年山东周边地区流行的H9N2 AIV是由不同进化方式而来;8 个基因节段关键氨基酸位点均发生了不同程度的变异,说明H9N2 AIV 在不断发生变异。因此,持续对山东周边H9N2 AIV分离鉴定和遗传进化分析,对我国家禽养殖、疫病防控以及公共卫生等方面均具有十分重要的意义。
猜你喜欢
铁道建筑技术(2022年10期)2022-10-28
中国典型病例大全(2022年12期)2022-05-13
中国典型病例大全(2022年9期)2022-04-19
复旦学报(医学版)(2021年5期)2021-10-13
养猪(2020年1期)2020-02-19
今日农业(2019年11期)2019-08-15
癌症进展(2018年11期)2018-12-30
国外畜牧学·猪与禽(2018年11期)2018-05-14
农村农业农民·B版(2015年9期)2015-10-16
医学研究杂志(2015年7期)2015-06-22
