
冷却治疗对劳力性热射病大鼠肾脏AQP2 mRNA及蛋白表达的影响
2019-11-19 12:31:10刘秀珍曹春梅张冰刘静民
中国运动医学杂志 2019年10期
刘秀珍 曹春梅 张冰 刘静民
1 清华大学体育部(北京100084)
2 巢湖学院体育学院(合肥238000)
劳力性热射病(exertional heat stroke,EHS)是一种危及生命的疾病,临床上表现为核心温度迅速升高超过40.5℃,伴有中枢神经系统功能异常及包括急性肾损伤在内的多器官功能障碍[1]。它对运动员生命安全造成极大的挑战,是马拉松等耐力运动员最常见的猝死原因[2]。劳力性热射病病情进展迅速,如果在30分钟内没有迅速得到降温处理,并发多器官功能障碍及导致死亡的机率会显著增加[3]。因此,热射病发作后,进行快速冷却治疗是确保热射病患者存活的重要手段[4]。Wohlfert 等的研究表明,冷水浸泡(cold water immersion,CWI)是降低热射病患者核心温度最快速的方法,可以使患者的存活率提升到接近100%[5]。
热射病病理生理学机制涉及热损伤、胃肠道炎症变化、凝血功能异常、水盐代谢失衡等方面,其中肾脏水盐代谢失衡与体温调节密切相关。目前国内外针对热射病的研究主要集中在热损伤、胃肠道炎症变化、凝血功能异常等方面,有关肾脏水盐代谢及热休克反应的研究很少。杜名的研究认为肾脏水通道蛋白与人类正常生理活动及多种疾病的发生发展密不可分,对水通道蛋白理论进行研究具有巨大的发展潜力[6]。但是,迄今为止,国内尚无冷却治疗对热射病患者肾脏水盐代谢调节方面的直接探索,冷却治疗在降温过程中,是否通过改变肾脏水通道蛋白(AQP2)的合成来改善水代谢,进而通过抗脱水途径恢复下丘脑体温调控作用并促进热射病病情转归的病理生理学机制需要进一步研究。因此,本研究针对劳力性热射病大鼠冷却治疗期间肾脏水通道蛋白基因转录中AQP2 mRNA及蛋白表达的变化进行研究,明确水通道蛋白在热射病水代谢稳态中的作用,为今后研究提供科学依据。
1 材料与方法
1.1 实验动物与分组
实验选用51 只7 周龄雄性Sprague-Dawley 大鼠,体重230~270 g,由北京市维通利华实验动物技术有限公司提供[SCXK(京)2016-0011]。大鼠分笼饲养,自由饮水摄食,室温20℃~25℃,湿度45%~50%,光照比12 h∶12 h。大鼠适应性喂养1 天,随机分为6 组,常温对照组(C,n=8)、高温运动即刻组(E,n=9)、高温运动休息1组(ER1,n=9)、高温运动冷浸1组(EC1,n=8)、高温运动休息2 组(ER2,n=9)和高温运动冷浸2 组(EC2,n=8)。
1.2 劳力性热射病大鼠模型建立与冷却治疗
1.2.1 适应性训练
参加高温运动的大鼠在常温下饲养,并在常温下进行为期5天的适应性训练。适应性训练跑台坡度为0。开始运动速度为18 m∕min,每天递增2 m∕min,最后达到26 m∕min;开始运动时间30 min,每天递增10 min,最后达到60 min。适应性训练安排见表1。

表1 大鼠适应性训练安排
1.2.2 劳力性热射病大鼠模型建立
适应性训练后休息3 天,在环境密闭的高温高湿实验室进行一次性高温力竭运动建立劳力性热射病大鼠模型。高温高湿实验室温度控制为36℃± 1℃,湿度控制为75% ± 5%。依据Bedford 最大摄氧量表[7]及高温运动大鼠预实验,确定大鼠高温运动时跑台坡度为5°,速度为28 m∕min,相当于60%~80%最大摄氧量的运动强度。依据(1)至(4)确定大鼠进行高温运动至力竭并使直肠温度上升到约42℃时为劳力性热射病建模成功:(1)Bouchama 等经典型热射病狒狒模型[8];(2)Lam 等经典型热射病大鼠模型[9];(3)Chang 等高温运动至力竭诱导劳力性热射病大鼠模型[10];(4)劳力性热射病经常在长时间耐力性运动(马拉松、越野赛及自行车公路赛等)中体温调节系统不堪重负时发生,在力竭或完赛的情况下发生率更高[11,12]。大鼠力竭标准为不能维持预定运动强度,在跑台后挡板滞留时,使用声、电刺激及毛刷连续驱赶3 次无效。采用北京搏贝科技有限公司JNT-200型鼠肛温计测量高温运动前及高温运动至力竭后大鼠直肠温度。测量方法为:将温度计探头插入大鼠直肠6.5 cm,固定探头,待读数稳定后读取数值。
1.2.3 冷水浸泡实验
依据(1)至(3)建立大鼠冷水浸泡方案:(1)McDermott等[13]治疗热射病常用冷却方式的冷却速率;(2)Riana 等[14]海军陆战队马拉松运动员劳力性热射病快速冷却流程;(3)Stewart 等[15]劳力性热射病患者冷却案例设定大鼠冷水浸泡预实验。冷水浸泡预实验选取19只EHS 大鼠,8 只死亡,11 只存活。结果如下:2 只2℃冰水浸泡4 s 的大鼠死亡;2 只9℃冷水浸泡10 min 的大鼠死亡;1 只19℃冷水浸泡10 min 的大鼠死亡;4 只2℃冰水浸泡2 s的大鼠,2只死亡,另2只存活;4只9℃冷水浸泡5 min的大鼠,1只死亡,3只存活;6只19℃冷水浸泡5 min的大鼠全部存活。为了确保EHS大鼠冷水浸泡实验的安全有效性,最终将本实验冷水浸泡温度设定为19℃,实际温度为18℃~20℃;冷水浸泡时间设定为5 min。大鼠休息温度设定为室温20℃,实际温度为19℃~21℃。
在高温运动至力竭后,ER1 组大鼠常温休息5 min;EC1 组大鼠冷浸5 min;ER2 组大鼠常温休息65 min(5 min+60 min);EC2 组大鼠冷浸5 min,常温休息60 min。冷浸或常温休息5 min 后,及常温休息60 min 后立即测量大鼠直肠温度。冷却速率用大鼠冷浸或休息5 min 期间单位时间内体温的减少量来表示,其计算公式为:
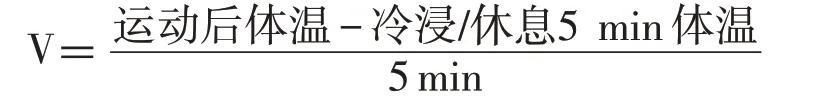
1.3 取材及处理
高温运动即刻组在高运动后即刻取材,常温对照组与高温运动即刻组同时取材。高温运动后,高温运动休息1 组、高温运动冷浸1 组、高温运动休息2 组和高温运动冷浸2组在完成相应的休息或冷浸实验后取材。采用10%水合氯醛按照0.32 ml∕100 g体重的剂量对大鼠进行腹腔麻醉。冰上解剖大鼠,选取腹主动脉取血;摘取肾脏放入液氮中迅速冷冻及冻存。
1.4 指标与测试方法
1.4.1 红细胞压积和血红蛋白
采用日本光电工业株式会社全自动血细胞分析仪(MEK-7222K,Japan)测试血常规,选取红细胞压积(hematocrit,Hct)和血红蛋白(hemoglobin,Hb)进行研究。相关试剂由上海东湖生物医学有限公司提供。
1.4.2 肾脏水通道蛋白AQP2 mRNA表达
采用荧光定量PCR 检测系统测试肾脏AQP2 mRNA 表达。取适量肾脏放入液氮预冷的研钵中研磨成粉末,转移到EP 管中;用Trizol(美国Invitrogen 公司)试剂抽提总RNA;用DNaseI 消化样品RNA 中的DNA;RNA 琼脂糖凝胶电泳;用反转录试剂盒(Takara)将RNA 反转录为cDNA;加入依据GenBank 数据库相关基因序列设计的特异性引物(见表3),加入EvaGreen 核酸染料Mix,以cDNA 为模板,在实时荧光定量PCR 仪(Roche LightCycler®480II)中进行Real Time-PCR 检测。分析产物溶解曲线并验证目标基因AQP2 和内参基因GAPDH 的扩增效率是否接近100%;根据各反应孔的目标基因与内参基因的△CT值,运用2-△△CT相对定量法计算目标基因AQP2 的相对表达量。
1.4.3 肾脏水通道蛋白AQP2蛋白表达

表2 实时荧光定量PCR引物信息
(1)蛋白抽提:预冷RIPA 蛋白抽提试剂,加入蛋白酶抑制剂(cocktail);4℃离心(12000 rpm×15 min)。取上清液。(2)BCA法蛋白定量:按照BCA蛋白定量试剂盒(北京康为世纪)使用说明操作,测定蛋白浓度。(3)Western Blot实验:取待检测蛋白样品进行电泳分离(浓缩胶恒压电泳:90 V,约20 min;分离胶恒压150 V,通过预染蛋白marker 来确定电泳停止时间)。采用湿转法将蛋白转至PVDF 膜,100 V 恒压,60 min。将膜浸没于5%BSA-TBST 中,水平摇床孵育1 h(RT)。一抗孵育:5%BSA-TBST 稀释一抗(AQP2,1∶500;GAPDH,1∶1000,Abcam公司),4℃水平摇床孵育过夜。次日,TBST洗膜:3×10min。二抗孵育:5%BSATBST 稀释二抗(山羊抗兔IgG(H+L),HRP,1∶10000,Abcam公司),室温孵育1 h。TBST洗膜:3×10 min。将ECL发光液滴加到膜的蛋白面,反应3~5 min。胶片曝光:10 s~5 min,显影2 min,定影。(4)计算结果:采用Image J软件计算蛋白条带灰度值,以目的蛋白与内参蛋白灰度值的比值作为目的蛋白相对表达量。
1.5 统计学方法
采用SPSS20.0 统计软件进行统计分析,结果以均值± 标准差(±s)表示。采用非参数检验(独立样本Mann-Whitney U检验)进行组间比较。采用非参数检验(相关样本Wilcoxon 带符号秩检验)对高温运动各组大鼠运动前与运动后直肠温度进行组内比较。P<0.05表示差异具有统计学意义。
2 结果
2.1 大鼠体重与运动时间
高温运动实验前,各组大鼠体重无显著性差异(P>0.05),见图1A。进行高温运动实验的5组大鼠运动时间无显著性差异(P>0.05),见图1B。
2.2 大鼠体温及冷却速率
表3显示,在高温运动实验前,与C组相比,E组大鼠体温显著升高(P<0.05);但是进行高温运动实验的5组大鼠体温相互间无显著性差异(P>0.05),提示适应性训练也会引起大鼠体温上升。
与运动前相比,E组、EC2组、ER1组与ER2组大鼠体温在高温运动后升高(P<0.05,P<0.01);EC1 组大鼠运动后体温升高,但无显著性差异(P>0.05)。
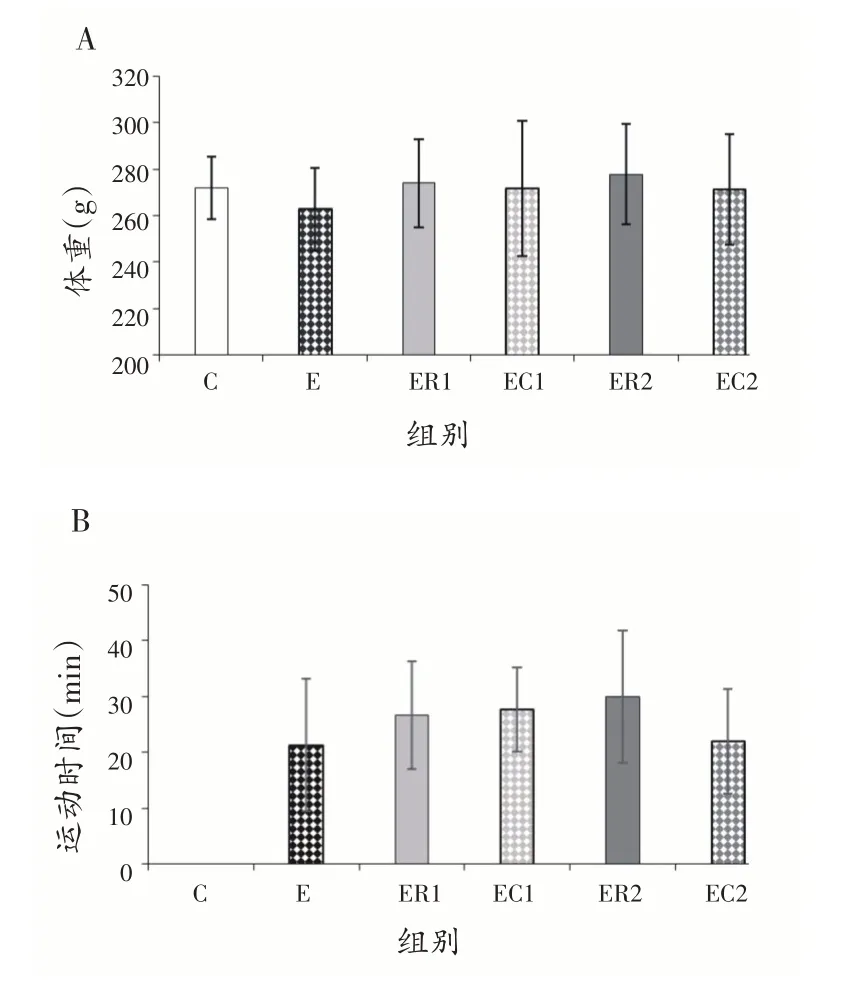
图1 各组大鼠的体重与运动时间
冷浸5 min 后,与ER1 组相比,EC1 组大鼠体温降低(P<0.01);与ER2 组相比,冷浸5 min 后的EC2 组大鼠体温降低(P<0.01),休息60 min 后的EC2 组大鼠体温降低(P<0.05)。与ER1 组相比,EC1 组大鼠冷却速率降低(P<0.01);与ER2 组相比,EC2 组大鼠冷却速率降低(P<0.01)。参加冷浸的EC1组和EC2组大鼠冷却速率无显著性差异(P>0.05)。
2.3 大鼠红细胞压积与血红蛋白
对各组大鼠的红细胞压积与血红蛋白进行分析,见表4。结果显示,与C组相比,E组大鼠红细胞压积升高(P<0.05),血红蛋白虽有升高,但无显著性差异(P>0.05);与E组相比,ER1组、EC1组、ER2组、EC2组大鼠红细胞压积及血红蛋白均无显著性差异(P>0.05);与ER1 组相比,EC1 组大鼠红细胞压积及血红蛋白均无显著性差异(P>0.05);与ER2 组相比,EC2 组大鼠红细胞压积及血红蛋白均无显著性差异(P>0.05)。
2.4 大鼠肾脏AQP2 mRNA表达
RNA 电泳图显示:提取的总RNA,经甲醛变性胶电泳观察,28S、18S、5S 三条带清晰易辨,可以证实RNA 的完整性,样品未发生降解,可以用于下游实验,见图2A。PCR 引物检测电泳图显示:根据电泳图中RNA 分子量标准,判断扩增产物长度大小与设计相符合;同时将扩增产物测序,测序结果证实引物扩增序列与目的基因匹配;说明引物可以用于荧光定量实验,见图2B。PCR 扩增曲线是样品PCR 反应实时荧光信号强度的反映,呈S 型,复孔的CT 值接近一致;反应完成后仪器会根据扩增曲线给出样品反应的CT值,用于相对表达量的计算,见图2C。PCR溶解曲线呈单峰,可以判断扩增产物单一,说明扩增产物的特异性,实验结果正常可用,见图2D。
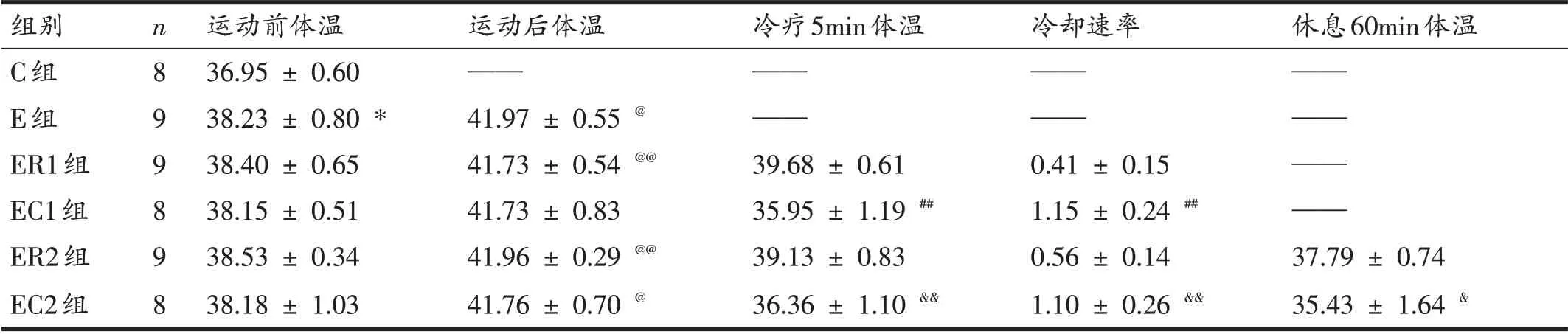
表3 各组大鼠的体温(℃)及冷却速率(℃∕min)
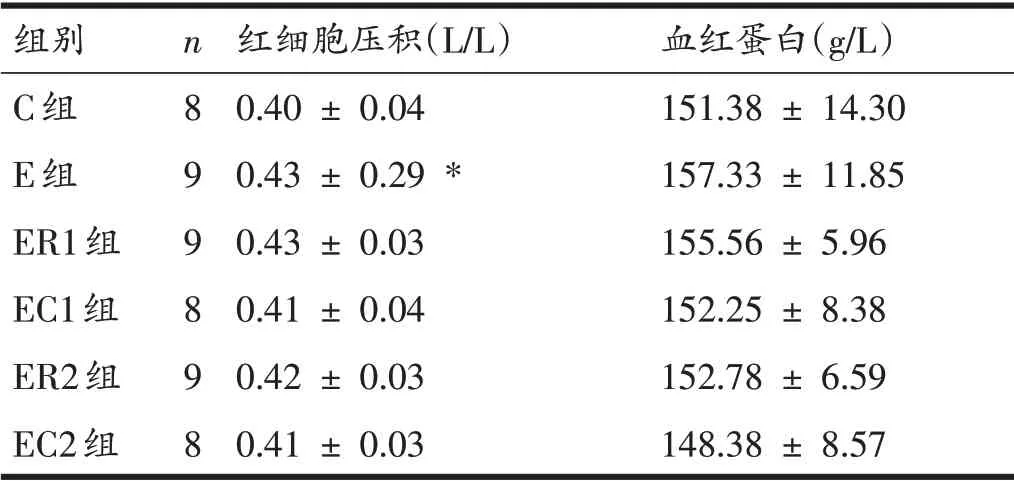
表4 各组大鼠的红细胞压积与血红蛋白
与C 组相比,E 组大鼠肾脏AQP2 mRNA 表达升高(P<0.01);与E组相比,常温休息ER1组和ER2组大鼠肾脏AQP2 mRNA 表达无显著性差异(P>0.05),经过冷浸的EC1 组和EC2 组大鼠肾脏AQP2 mRNA 表达降低(P<0.01);与休息ER1 组相比,冷浸EC1 组大鼠肾脏AQP2 mRNA 表达降低(P<0.01);与ER2 组相比,EC2 组大鼠肾脏AQP2 mRNA 表达降低(P<0.05),见图2E。
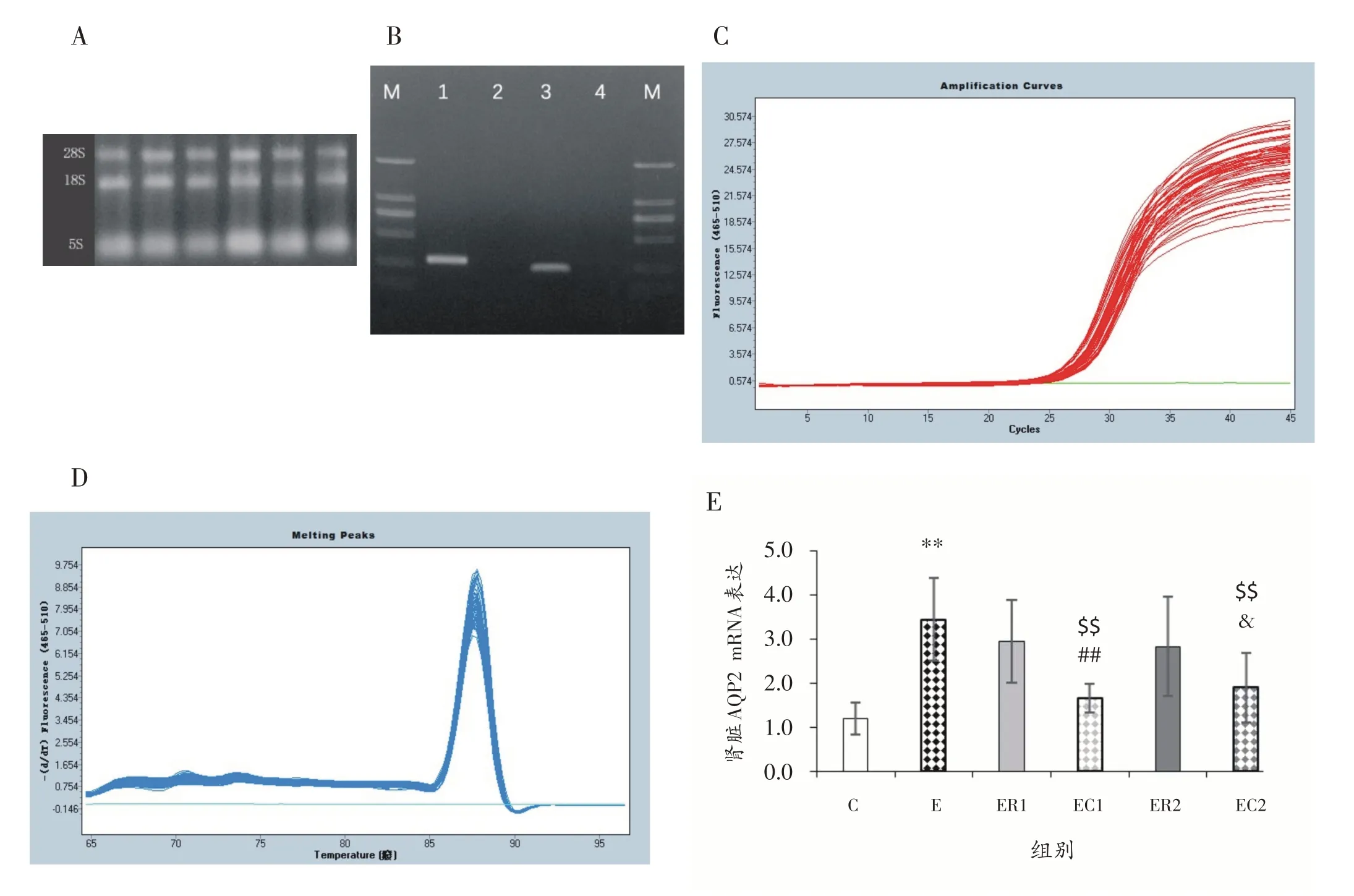
图2 RNA电泳和PCR结果
2.5 大鼠肾脏AQP2蛋白表达
图3显示,与C组相比,E组大鼠肾脏AQP2蛋白表达升高(P<0.01);与E 组相比,常温休息ER1 组和ER2组大鼠肾脏AQP2蛋白表达无显著性差异(P>0.05),经过冷浸的EC1 组和EC2 组大鼠肾脏AQP2 蛋白表达降低(P<0.05);与ER1 组相比,EC1 组大鼠肾脏AQP2 蛋白表达无显著性差异(P>0.05);与ER2 组相比,EC2 组大鼠肾脏AQP2蛋白表达无显著性差异(P>0.05)。
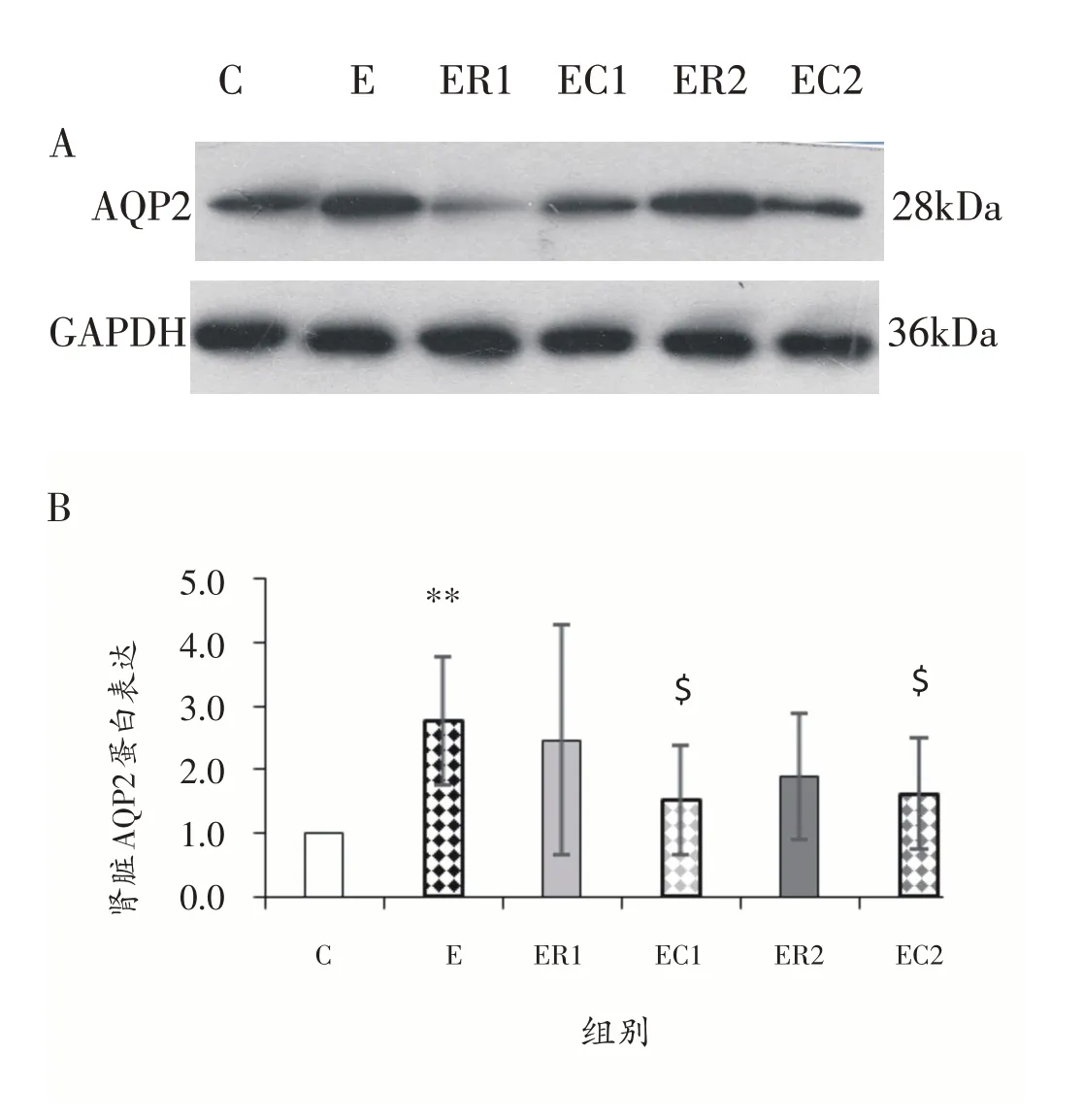
图3 Western Blot结果
3 讨论
3.1 大鼠体温变化
热射病动物模型分为两大类:(1)在高温环境下进行热暴露建立的热射病动物模型称为经典型热射病动物模型;(2)在高温环境下进行热运动建立的热射病动物模型称为劳力性热射病动物模型。
在实验室劳力性热射病动物模型的研究中,Zhang等以45℃高温环境下热运动至力竭作为热射病诊断标准,建立了劳力性热射病大鼠模型,用于研究红景天苷对线粒体的保护作用,大鼠在建模前并未进行适应性训练[16]。为了研究劳力性与经典型热射病大鼠模型体温调节差异及预后特征,帅军等安排大鼠在39℃高温环境中运动,并以核心温度升高到42℃作为热射病诊断标准,建立了劳力性热射病大鼠模型,所有大鼠均进行6 天适应性递增负荷跑台训练后再进行分组,适应性训练是否会引起大鼠体温变化未见报道[17]。
与前人的研究结果有所不同,本研究中的常温对照组未进行适应性训练,但是5 个高温运动组均进行了适应性训练,实验结果提示适应性训练也会引起大鼠体温上升。本研究中,在36℃高温环境下热运动至力竭的5组大鼠在运动后体温均上升到42.0℃左右,符合劳力性热射病大鼠建模标准,说明劳力性热射病大鼠建模成功。
快速降温是热射病救治的关键。McDermott 等的研究认为,在诸多降温措施中,冷水浸泡的冷却速率最快,研究发现2℃冷水浸泡时冷却速率是0.35℃∕min,8℃和20℃冷水浸泡时冷却速率均为0.19℃∕min。另外一些冷却策略,如冷却水雾喷洒、冷却液体静脉注射、冰敷外周动脉等,由于冷却速率过低而不建议采用[13]。在冷却治疗时间的设置上,Stewart 等的研究认为劳力性热射病患者通常具有41.0℃~43.5℃的核心体温以及38.9℃的冷却终止点体温,依据每5 min 降低1℃的冷却速率,提出冷水浸泡15 min或者提供合理的冷却终止点进行冷却治疗[15]。冷水浸泡时的冷却速率存在运动项目差异。Godek等的研究表明,越野赛运动员在10℃冷水浸泡时的冷却速率为0.255℃∕min,核心温度从39.5℃降到37.5℃仅需要7.7 min;足球运动员的冷却速率0.156℃∕min,核心温度从39.5℃降到37.5℃需要11.4 min[18]。本研究中两组大鼠冷水浸泡时冷却速率分别为1.15℃∕min 和1.10℃∕min,比人体降温速率更快。两组大鼠冷浸5 min 后体温均降到低于运动前体温,说明冷浸5 min 对劳力性热射病大鼠体温已产生显著影响。与人体相比,大鼠在冷浸时的冷却速率更快,冷浸时间更短,提示物种或体重差异可能会产生不同的热调节反应。
3.2 大鼠血液浓缩程度
血常规检查中红细胞压积和血红蛋白是反映血液浓缩程度的重要指标。剧烈运动时大量出汗会引起机体脱水,血液浓缩,红细胞压积和血红蛋白升高。全军重症医学专业委员会于2015年制定热射病规范化诊断与治疗专家共识(草案),提出劳力性热射病发病早期因脱水致血液浓缩可出现血红蛋白(Hb)升高、红细胞压积(Hct)增加、血小板(PLT)发病初期正常继而迅速下降等血常规异常症状[19]。与前人的研究相一致,本研究中E组大鼠红细胞压积显著高于C组大鼠,说明大鼠进行高温运动确实会因为脱水而引起机体血液浓缩。与C 组大鼠相比,E 组大鼠血红蛋白浓度虽然升高,但差异并没有统计学意义,提示血红蛋白在反映血液浓缩程度方面不够敏感或可能存在滞后现象。本研究中,E组大鼠红细胞压积显著性升高,提示红细胞压积是血液浓缩程度的敏感指标,可作为劳力性热射病发病的病理生理学标志物。
在针对劳力性热射病从运动现场到急诊部的处理流程中,Riana提出在运动现场进行冷水浸泡是热射病患者救治的首选方案;随后应进行实验室检测,包括钠(Na)、钾(K)、氯(Cl)、葡萄糖(Gluc)、肌酐(Cr)、血尿素氮(BUN)和红细胞压积(Hct)等;治疗脱水、低血糖和低钠血症等[14]。关于冷却对血液浓缩程度影响的研究鲜有报道。在研究大鼠血液粘度对温度的依赖性时,Lipina等发现冷却会使大鼠血液浓缩,红细胞压积升高[20]。与Lipina等的研究有所不同,本研究中冷浸虽然引起大鼠红细胞压积及血红蛋白下降,但并未发生显著性变化,提示红细胞压积和血红蛋白对冷水浸泡带来的温度变化不够敏感或可能存在滞后现象。引起红细胞压积改变的根本原因是脱水,而不是温度。冷浸对红细胞压积没有明显影响,其原因可能是冷却速率过快及冷浸时间较短(只有5 min),以至于机体的生化过程来不及作出反应,未能明显加重脱水(利尿途径)或缓解脱水(抗利尿途径),因而未能引起红细胞压积发生显著性变化。
3.3 劳力性热射病大鼠冷却治疗中水通道蛋白调节机制探讨
Hasegawa 等的研究发现,运动时水分丢失是引起体温升高的重要因素[21]。机体水代谢主要在肾脏由水通道蛋白进行调控。AQPs 是一类具有跨膜水转运功能的蛋白质家族。目前在肾脏组织共发现了8 种AQPs,其中AQP1-4 在调节水转运中起关键作用[22]。AQP2主要分布于集合管顶端质膜,是响应抗利尿激素(亦称精氨酸加压素,AVP)调节集合管对水通透性的主要靶分子,在维持水平衡中起重要作用。
为了探索高温环境下力竭运动对大鼠肾脏水通道蛋白AQP2 的影响,董柔等进行了相关的研究,发现在38℃环境下以20 m∕min 的速度运动至力竭的大鼠AQP2 mRNA 表达非常显著性上调,AQP2 蛋白表达上调,但未引起显著性变化。该研究认为高温力竭运动可通过上调AQP2 mRNA 表达增强肾脏对水的重吸收,AQP2蛋白表达上调不显著,可能与AQP2蛋白表达滞后于AQP2 mRNA 表达有关[23]。但该研究并未涉及热射病对AQP2 mRNA表达及AQP2蛋白表达的影响。
水通道蛋白与人类健康密不可分,是疾病的特殊标志物。据报道,在包括心力衰竭和肝硬化在内的各种水排泄受损的疾病中,AQP2 在肾脏的含量会改变,AQP2的尿排泄增加[24]。热射病实验室检查中,有关水通道蛋白的研究并不常见。Du 等研究了热射病尸检人脑标本中大脑皮层基质金属蛋白酶(matrix metalloproteinases,MMPs)、紧 密 连 接 蛋 白(claudin5,CLDN5)、封闭蛋白(occludin,OCLN)、闭锁小带蛋白(zona occludens protein-1,ZO1)和水通道蛋白(aquaporins,AQPs)表达增加与热射病脑水肿的关系,发现热射病患者MMP9 mRNA、OCLN mRNA、ZO1 mRNA、CLDN5 mRNA、AQP4 mRNA显著性升高,且与脑含水量增加呈正相关,但并未进行AQP2的研究[25]。
水通道蛋白调节机体水代谢。热射病诱导的肠道通透性改变可能与水通道蛋白对机体水代谢的调节有关。Wang 等研究了包括AQP2 在内的10 种水通道蛋白在热射病大鼠小肠组织中不同恢复时间点的表达情况,发现热射病发作时AQP7、AQP8 和AQP11 mRNA表达显著性上调,热射病后恢复1小时AQP1、AQP3和AQP5 mRNA表达显著下调,提出热射病诱导小肠损伤和细胞凋亡与AQPs 上调∕下调时水转运改变密切相关,但此研究中并未在小肠检测到包括AQP2、AQP4、AQP6和AQP9在内的4支基因的mRNA表达[26]。
与前人的研究有所不同,本研究主要聚焦在肾脏集合管水代谢。本研究中热射病大鼠(E组)与常温对照组(C 组)相比,水通道蛋白AQP2 mRNA 表达和AQP2蛋白表达均出现上调(P<0.01),说明高温运动引起劳力性热射病的同时导致AQP2合成显著性增加,机体对水分的重吸收增强,以对抗机体的脱水情况。提示AQP2 mRNA 表达和AQP2 蛋白表达是劳力性热射病病理生理学改变的敏感指标,可作为劳力性热射病发病的病理生理学标志物。
有关低温对水通道蛋白影响的研究很少报道。水通道蛋白调节细胞质膜间微妙的渗透平衡,渗透损伤是冷冻过程中引起细胞死亡的主要因素。水通道蛋白AQP2 上调和转运会减轻低温冷藏过程中冷诱导渗透损伤。Wang 等的研究表明,在冷冻前用抗利尿激素(AVP)处理大鼠肾髓质集合管(inner medullar collecting duct,IMCD)细胞,使集合管细胞外质膜上的AQP2 含量增加,可以减弱冷诱导(-4℃)引起的大鼠肾髓质集合管细胞渗透损伤[27]。本研究中,经过冷浸的EC1 组和EC2 组大鼠与E 组大鼠相比,肾脏AQP2 mRNA 表达非常显著性下调;AQP2 蛋白表达显著下调。EC1组大鼠与ER1组大鼠相比,肾脏AQP2 mRNA表达非常显著性下调;AQP2 蛋白表达无显著性差异。EC2组大鼠与ER2组大鼠相比,肾脏AQP2 mRNA表达显著性下调;AQP2蛋白表达无显著性差异。这种现象说明冷浸对肾脏AQP2 mRNA 表达的影响最显著,对AQP2蛋白表达的影响次之;进一步证明冷浸影响肾脏水通道蛋白的合成,且这种影响符合从肾脏AQP2 mRNA表达到AQP2蛋白表达的逐级滞后的合成顺序;肾脏AQP2 mRNA 表达和AQP2 蛋白表达对冷浸带来的温度变化敏感,其机制可能与AQP2在低温下通过抑制抗脱水机制重建自身体温调节有关,有待进一步的研究进行证明。
4 结论
冷浸可以有效降低热射病大鼠机体核心温度;冷浸诱导水通道蛋白AQP2 mRNA 表达及AQP2 蛋白表达显著性下调,导致水通道蛋白在肾脏的合成减弱;冷浸对红细胞压积Hct及血红蛋白Hb没有明显影响。
猜你喜欢
中国实用医药(2024年8期)2024-05-29 03:43:58
昆明医科大学学报(2021年5期)2021-07-22 07:32:32
解放军医学院学报(2020年12期)2020-03-29 05:11:38
解放军医学院学报(2020年12期)2020-03-29 05:11:26
当代旅游(2019年1期)2019-08-30 03:37:10
今日农业(2019年12期)2019-08-15 00:56:32
中国兽医杂志(2018年3期)2018-06-11 09:36:32
教师·上(2016年6期)2016-12-01 09:20:37
兽医导刊(2016年12期)2016-05-17 03:51:33
河南医学研究(2015年11期)2015-03-28 00:46:42
