
Tribbles同源蛋白3与巨噬细胞极化和胰岛素抵抗的关系及运动对其调控作用的研究进展
2019-11-19 12:31:12罗维艾磊李显王博发周越
中国运动医学杂志 2019年10期
罗维 艾磊 李显 王博发 周越
1 南京体育学院运动健康科学系(南京210014)
2 北京体育大学运动生理学教研室(北京100084)
3 江苏省体育科学研究所(南京210033)
糖尿病是继癌症和心血管疾病后威胁人类健康的第三大慢性疾病[1],其中2 型糖尿病(type 2 diabetes mellitus,T2DM)患者占比高达90%[2,3]。作为T2DM的典型病理特征,胰岛素抵抗是T2DM 的发病基础,更导致T2DM 的进一步发展[4]。无论T2DM 病因如何,最终都表现出机体产生明显的胰岛素抵抗而发生病变[5]。近些年慢性炎症被认为是多种慢性疾病的发病基础,其在胰岛素抵抗发生发展中的作用受到广泛关注。巨噬细胞(macrophages)作为重要炎症细胞,可在不同应激诱导下发生极化,在机体产生促炎或者抗炎的作用,进而影响机体胰岛素敏感性[6]。作为21世纪新发现的假激酶,Tribbles 同源蛋白3(Tribbles homology protein 3,TRIB3)具有广泛生物学活性,是目前寻找癌症、心血管疾病、糖尿病等慢性疾病药物靶点的研究热点[7],同时因其在运动应激下产生适应性变化而在运动科学研究领域引起关注。国内外研究表明TRIB3与巨噬细胞极化、胰岛素抵抗和运动训练均存在密切联系。
1 TRIB3的结构与功能
Tribbles 是最早在果蝇体内被发现的一类假性蛋白激酶家族,该家族成员的激活均可降解细胞周期调控因子,抑制细胞有丝分裂。迄今发现的人源性Tribbles 基因有三个,Tribbles homology protein1(TRIB1),Tribbles homology protein2(TRIB2)和TRIB3。作为被确认的癌症基因,TRIB1和TRIB2在癌症疾病的发生发展中被充分研究。TRIB3是抑制细胞有丝分裂并诱导细胞凋亡的核基因,是近些年国内外研究的热点分子[7],其主要由3 个功能区构成:中心假激酶区,N 末端和C末端。C末端包含两个保守序列,分别为ILLHPW模体构成的丝裂原活化蛋白激酶激酶(mitogen-activated protein kinase kinase,MAPK 激酶)MEKs 结构域和由DQLVP 模体构成的E3 泛素连接酶COP1 结构域[8,9]。MEKs 结构域为MAPK 激酶的结合位点,通过靶向MEKs 家族来特异性地调节MAPK 家族磷酸化水平,介导TRIB3与多种MAPK间的相互作用[10,11]。COP1结构域主要调节TRIB3蛋白的泛素化作用。
现有研究显示,TRIB3 可被多种应激因素诱导上调表达,是重要应激基因,同时其又具有广泛生物学活性[12]。内质网应激、低氧、营养过量、营养缺乏等均可上调或下调TRIB3表达水平[13],通过与不同激酶依赖性蛋白质相结合参与转化生长因子-β(transforming growth factor-β,TGF-β)、磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)、MAPK、核因子κB(nuclear factor kappa-B,NF-κB)等多种信号通路的调节,在炎症、血糖和脂质代谢、脂肪细胞分化、细胞凋亡等过程中发挥重要作用[14,15]。例如,Liu 等研究发现小鼠禁食后脂肪中TRIB3 表达增加,并通过其C 末端直接结合E3 泛素连接酶,促进乙酰辅酶A 羧化酶1(acetyl-CoA carboxylase 1,ACC1)的泛素化与降解,进而诱导脂肪细胞分解[16]。Schisler等研究发现小鼠脂肪组织中过表达TRIB3后,其脂肪酸氧化能力得到提高,且被保护免受高脂饮食诱导的肥胖[17]。另外,大量研究发现TRIB3 对应激的响应具有组织特异性,即同一应激条件下不同组织中TRIB3 产生不同响应,以精确调节机体应激和代谢的动态平衡[18]。例如Liu 等研究发现禁食条件下TRIB3 在脂肪组织上调,而在肌肉组织出现下调;营养过剩条件下TRIB3在脂肪组织下调,而在肌肉组织出现上调,即在营养缺乏时促进能量从脂肪到骨骼肌转移,维持骨骼肌机能正常;在营养过剩时促进能量从骨骼肌到脂肪的转移,减轻骨骼肌负担的同时促进能量在脂肪组织储备[16]。
2 TRIB3在巨噬细胞极化中的作用
炎症是机体固有的保护性免疫机制,但慢性炎症的发生会诱导疾病,是癌症、心血管疾病与糖尿病等慢性疾病的发病基础[19]。近些年来慢性炎症在胰岛素抵抗发生发展中的作用广受关注。作为重要的炎症细胞,巨噬细胞在机体免疫机能的调节中至关重要。在组织不同微环境影响下,巨噬细胞分泌的趋化因子和细胞因子、对趋化因子的应答以及吞噬能力等都会发生改变,形成功能迥然的经典极化M1型和替代性极化M2型[6],即产生巨噬细胞极化。M1型巨噬细胞主要分泌白细胞介素-1(interleukin-1,IL-1)、白细胞介素-6(interleukin-6,IL-6)、肿瘤坏死因子-α(tumor Necrosis Factor-α,TNF-α)等炎症因子,增强炎症反应。M2型巨噬细胞主要分泌白细胞介素-10(interleukin-10,IL-10)等抗炎因子,抑制炎症反应,在机体产生抗炎效应[20]。作为与细胞凋亡相关的基因,TRIB3表达上调抑制巨噬细胞细胞因子的表达且细胞凋亡增加,而敲低TRIB3则可抵抗巨噬细胞凋亡[7]。
已有研究显示TRIB3作为炎症信号转导的调节因子参与多种关键炎症信号通路的调节,包括MAPK 通路和PI3K通路,在不同种类细胞中产生不同的调控作用[14],Zhang 等研究发现TRIB3 通过调节MAPK 家族成员ERK1∕2 及其下游通路,增加脂肪组织中脂肪酸氧化,在营养过剩的脂肪组织中产生抗炎作用[14]。关于TRIB3对巨噬细胞极化的影响,有研究发现TRIB3表达增加时,由M1型巨噬细胞分泌的炎性细胞因子,如IL-1β、IL-3 和IL-6 等表达增加[21]。地塞米松和cAMP 等抗炎药通过转录因子CREB 和叉头转录因子1(forkhead box O1,FoxO1)可降低TRB3的表达,瘦素和低密度脂蛋白等脂肪因子可增加TRIB3的表达[22,23]。因此,TRIB3 与巨噬细胞极化和炎症因子相互调节,在疾病发展中发挥重要作用,有望成为慢性炎症性疾病的潜在治疗靶点。
3 TRIB3在胰岛素抵抗中的作用
3.1 胰岛素抵抗与骨骼肌萎缩
胰岛素抵抗是T2DM 的主要病征,表现为一定浓度胰岛素产生的效应降低,实质上是胰岛素激活的葡萄糖转运和骨骼肌等靶器官的葡萄糖代谢能力的降低,在分子水平上是胰岛素的信号转导通路受损所致[24]。除了糖尿病,胰岛素抵抗还是肥胖、动脉粥样硬化以及高血压等常见的慢性代谢性疾病发病的共同诱因[4]。作为胰岛素重要的靶器官,骨骼肌在胰岛素抵抗发生发展中扮演重要角色。胰岛素刺激下人体80%以上的葡萄糖摄取在骨骼肌中完成,而骨骼肌质量减少是包括糖尿病、癌症、心衰等在内的多种慢性疾病的共同特征[25],也是糖尿病易感的早期预测指标[26],且减少的趋势随糖尿病病程发展而加剧[27],最终导致骨骼肌萎缩[28]。骨骼肌萎缩导致外周血糖摄取能力减弱,进一步加快糖尿病发展进程,进入糖尿病→肌萎缩→糖尿病的恶性循环[29]。同时,骨骼肌质量丢失及力量下降也严重影响生活质量[30]。
研究表明,骨骼肌细胞存在胰岛素信号转导和骨骼肌蛋白合成的共同途径,导致糖尿病与骨骼肌萎缩间的恶性循环[31]。PI3K∕蛋白激酶B(protein kinase B,AKT)通路将多种分子信号从细胞外传递至细胞内,调节糖转运及细胞增殖分化等重要生理过程[32]。在胰岛素信号转导中,胰岛素刺激下胰岛素受体底物1(insulin receptor substrate1,IRS1)∕PI3K∕AKT 通路激活,介导重要的糖摄取分子葡萄糖转运子4(glucosetransporters 4,GLUT4)转运至细胞膜,将血液中葡萄糖摄取入细胞内,而胰岛素抵抗的发生通常表现为该通路受损[33]。在骨骼肌蛋白质合成中,PI3K∕AKT 通路的激活下调骨骼肌萎缩的关键因子萎缩相关基因-1(atrophy gene-1,Atrogin-1)与肌肉特异性环指蛋白1(muscle specific RING finger protein 1,MuRF1)转录水平,抑制蛋白降解并促进蛋白合成。胰岛素抵抗和∕或肌萎缩发生时,IRS1∕PI3K∕AKT 途径受阻并上调Atrogin-1和MuRF1 基因表达,引发蛋白酶体降解[34]。大量研究发现,胰岛素抵抗患者AKT活性降低,FoxO3水平升高,Atrogin-1、MuRF-1 均出现上调[35],提示细胞内AKT 活性降低,Atrogin-1与MuRF1的激活是胰岛素抵抗和骨骼肌萎缩的共同原因,IRS1∕PI3K∕AKT 通路是潜在通路。因此,IRS1∕PI3K∕AKT通路激活水平及其是否受损可较好地反映机体胰岛素抵抗和骨骼肌萎缩情况及二者间的相互作用,如Li等检测IRS1∕PI3K∕AKT通路蛋白激活水平作为体外胰岛素抵抗模型的建模评价标准[36]。
3.2 TRIB3在胰岛素抵抗中的重要作用
研究证实TRIB3在肝脏、胰岛、脂肪等不同组织中表达上调会损伤胰岛素诱导的糖摄取,而下调则提高胰岛素敏感性[18]。Örd 等通过研究提出禁食应激诱导肝细胞TRIB3 激活增加,并抑制AKT 不同位点磷酸化激活,最终导致肝糖原合成下降,糖异生增多,血糖水平升高,诱发胰岛素抵抗[37]。Bi 等研究发现大量酒精摄入可增加肝脏TRIB3 的表达,并导致胰岛素抵抗的发生[38]。Qian 等研究表明胰岛β细胞中TRIB3 表达上调会促进细胞凋亡并加重糖尿病[39]。Liu等研究发现禁食条件下TRIB3 在脂肪组织表达上调,机体血糖水平升高[16]。而关于TRIB3 对高糖高脂应激下骨骼肌细胞的影响则存在争议,大部分研究认为高糖高脂应激下,骨骼肌中TRIB3 上调,诱导骨骼肌细胞中胰岛素抵抗的发生发展,并指出TRIB3 是骨骼肌能量代谢的关键调节因子,是营养过剩条件下诱发胰岛素抵抗的必要参与因子,提出抑制TRIB3 可逆转糖尿病中的胰岛素抵抗和减少“葡萄糖毒性”,可作为治疗糖尿病的新靶点[40]。但也有研究提出TRIB3 并不是导致胰岛素信号转导通路受损和胰岛素抵抗的必要基因,例如Nicolas等敲除胰岛素抵抗应激下骨骼肌细胞中的TRIB3基因后,AKT磷酸化水平仍被抑制,即TRIB3敲除无法逆转胰岛素抵抗诱导的一系列生理病理效应[41]。Ding 等研究发现小鼠骨骼肌组织中过表达TRIB3 后,其胰岛素信号转导、肌肉葡萄糖摄取和全身血糖稳态并未受影响[42]。
4 巨噬细胞极化与胰岛素抵抗的关系
代谢应激下,巨噬细胞越来越多的渗入骨骼肌,参与骨骼肌代谢调节[43]。诸多研究证实巨噬细胞在胰岛素抵抗的发生发展中起到关键作用,巨噬细胞M1型和胰岛素抵抗的发生发展相关,M2型和胰岛素敏感性的提高相关,且巨噬细胞与胰岛素靶细胞-骨骼肌细胞间存在交叉对话,进一步调节着巨噬细胞在胰岛素抵抗发生发展中的作用[44]。离体和在体研究都发现葡萄糖、棕榈酸及胰岛素混合物可诱导巨噬细胞分化为M1型促炎巨噬细胞[45]。Yuan等研究发现肥胖诱导脂肪组织中巨噬细胞向M1型极化增加,向M2型极化减少,导致巨噬细胞更多地向促炎表型发展并诱发胰岛素抵抗。特异性的在骨骼肌中过表达IL-10 可减少由于高脂应激导致的巨噬细胞浸润与炎症反应,最终达到提高胰岛素敏感性的效果[46],且巨噬细胞M2 型是IL-10等抗炎因子的主要来源。Wan等将骨骼肌细胞与巨噬细胞共培养,同样发现M1型巨噬细胞通过产生更多促炎因子来降低骨骼肌细胞胰岛素敏感性,导致糖耐量受损和胰岛素抵抗的发生[47]。
虽然在高糖高脂等应激下,巨噬细胞极化参与机体代谢调节,影响如糖尿病、肥胖等代谢性疾病的发生和发展,但其具体调控机制尚不清楚[48]。深入研究高糖高脂应激诱导巨噬细胞极化的具体机制以及巨噬细胞与主要胰岛素靶细胞-骨骼肌细胞间的交互作用对于寻求胰岛素抵抗的病因,探索防治糖尿病、高血压等慢性代谢疾病的有效手段具有重要意义。TRIB3与巨噬细胞极化和胰岛素抵抗关系密切,是否可将TRIB3作为靶点研究高糖高脂应激对巨噬细胞极化和胰岛素抵抗的影响,值得深思。
5 TRIB3在运动训练中的适应性改变
运动在慢性疾病的防控和骨骼肌表型重塑中发挥着重要作用[49],阐明运动影响机体代谢与骨骼肌表型重塑的分子机制至关重要[50]。关于运动对TRIB3 调控的研究,主要集中在一次急性运动或长期有氧运动对TRIB3及其下游通路的影响,且存在争议。
大部分研究支持一次急性运动可以减少TRIB3表达,如Flowers 等研究发现游泳运动后饮食诱导的肥胖小鼠肝组织中TRIB3 的表达与静息小鼠相比降低[51]。Rodrigues等提出一次长时间游泳运动后高脂饮食的肥胖大鼠下丘脑TRIB3 蛋白表达降低,促进AKT 磷酸化激活,提高胰岛素敏感性[52]。Henrique等通过研究一次长时间跑台运动对高脂饮食诱导的肥胖小鼠的影响,同样发现高脂饮食后小鼠TRIB3 表达增加,而运动后TRIB3 表达比运动前下降59%,且与AKT 磷酸化水平呈负相关[53]。Moyas 等也指出急性运动8 小时后,与安静组相比,运动组TRIB3表达明显降低[54]。
虽然大部分研究支持运动降低TRIB3 表达,但也有研究提出运动对TRIB3 表达无明显影响,如Gaspar等提出一次急性跑台运动后大鼠骨骼肌TRIB3表达未见明显差异[55]。由于肥胖诱发的内质网应激是TRIB3表达增加的主要诱因,而该研究所使用的动物模型未出现肥胖,这可能是导致该研究中运动对TRIB3 表达未产生明显影响的原因。Ding等提出TRIB3过表达小鼠骨骼肌纤维类型向氧化型I∕IIa 型转变,且运动能力得到提高,未出现胰岛素抵抗[42],产生该矛盾结果的原因可能是该研究模型以TRIB3过表达转基因鼠作为研究对象,TRIB3 在小鼠体内出现长期慢性的高表达导致胰岛素功能出现代偿,且该研究中动物模型均为正常饮食,也未产生肥胖,若以高脂饮食干预再进行运动,效果不得而知。另外,TRIB3过表达提高运动能力可能与TRIB3调节运动中能量代谢的功能有关。
另外,虽目前未见长期离心运动(如下坡跑台运动、下楼梯等)与TRIB3 间关系的研究,但现有研究表明一次长时间离心运动后骨骼肌结构产生损伤性变化,并伴随炎症反应[56],而重复性离心运动并不会加重肌肉损伤和炎症反应,且能上调生肌因子5(myogenic factor5,Myf5)和成肌调节因子(myogenin)等肌再生因子表达[57],说明离心运动通过调节机体炎症水平来促进骨骼肌肌纤维重塑,改善机体胰岛素抵抗。TRIB3参与机体多种关键炎症信号通路的调节,离心运动是否是通过调节TRIB3 的表达来调节机体炎症水平,改善机体胰岛素抵抗、促进骨骼肌重塑,还有待进一步深入研究。
6 小结
TRIB3 在不同应激下表达存在组织特异性,且目前关于TRIB3 与运动训练关系的研究结果存在争议,综合当前大部分的研究结果,高糖高脂等刺激下骨骼肌TRIB3表达上调,进而抑制MAPK和AKT信号激活,并促进NF-κB 表达增加,诱发巨噬细胞M1 型极化,产生炎症,同时导致骨骼肌蛋白降解增加,骨骼肌质量下降,二者交互对话共同诱发胰岛素抵抗并导致胰岛素抵抗与骨骼肌萎缩间恶性循环,运动可下调TRIB3 表达,促进AKT磷酸化激活,可能在改善巨噬细胞极化状态和胰岛素抵抗中发挥作用(图1)。
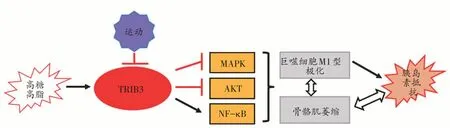
图1 TRIB3与骨骼肌炎症、胰岛素抵抗及运动训练的关联
慢性炎症是肥胖、动脉粥样硬化和糖尿病等慢性代谢性疾病发生发展的核心,巨噬细胞作为主要炎症细胞,通过不同极化分型参与机体代谢调节,影响着糖尿病等多种代谢性疾病的发生发展。提高骨骼肌质量对于改善胰岛素抵抗、预防和控制糖尿病等多种慢性代谢性疾病的发生发展具有重要意义。TRIB3被高糖高脂等多种应激诱导激活,且具有广泛生物学活性,在炎症、胰岛素抵抗中都发挥着重要作用。运动在调节机体炎症水平和骨骼肌重塑中发挥着重要作用,且运动训练可调节TRIB3 的表达。以TRIB3 为靶点,探索高糖高脂应激下巨噬细胞极化和胰岛素抵抗的调控机制及其在运动训练中的适应性变化或许可作为糖尿病等慢性代谢综合征的治疗和控制研究切入点。
猜你喜欢
中老年保健(2021年5期)2021-08-24 07:06:20
小雪花·成长指南(2021年2期)2021-05-20 09:14:00
中华养生保健(2020年5期)2020-11-16 01:44:36
天津医科大学学报(2019年3期)2019-08-13 06:53:00
初中生世界·九年级(2019年4期)2019-05-05 01:07:12
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
医学研究杂志(2015年12期)2015-06-10 06:57:46
医学研究杂志(2015年5期)2015-06-10 06:43:26
