
高温冲击对受丝茅入侵的细叶结缕草现实和潜在竞争力的影响
2019-08-30 02:26:52汪智宇李莹刘金平伍德苟蓉
草业学报 2019年8期
汪智宇,李莹,刘金平,伍德,苟蓉
(西华师范大学西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009)
四川盆地为典型的亚热带湿润季风气候,年均气温17.4 ℃,年有霜期仅13.7 d,以前常用高羊茅(Festucaarundinacea)、草地早熟禾(Poapratensis)、黑麦草(Loliumperenne)建植单一或混播冷季型草坪,以期实现四季常绿的绿化效果[1]。该区夏季≥30 ℃约120 d,且高于40 ℃的酷热天气频现[2],而冷季型草坪适宜生长温度为15~25 ℃[3],长时间高温、高湿致使冷季型草坪分生再生能力差、夏眠期长、病害频发,管护疏忽极易使建成草坪1~3年退化殆尽[4]。近年来,从国内外引进数种结缕草属(Zoysia)草种建植暖季型草坪,其中具有低矮、纤细、节间短等生物学特征和耐贫瘠、耐践踏、抗干旱、耐盐土、耐低修剪[5]等生态学特性的细叶结缕草(Z.tenuifolia),在该区观赏性草坪、运动场草坪和设施草坪建植中得到广泛应用。
草坪植物长期经低修剪、高水肥等维持景观的养护后,植株低矮纤细、茎叶幼嫩多汁、根系浅表化,其拓展空间、吸收资源和抵抗危害能力下降,极易被杂草入侵和危害,故防除杂草是草坪养护最重要和最基本的工作,除杂用工占总用工的80%左右,费用占养护费用的60%左右。细叶结缕草具根茎和匍匐茎,拓展能力和分生再生能力强[6],在25~35 ℃的正常养护下,生长旺盛,极易形成低矮、致密草层[7],凭借强大匍匐茎和根茎形成致密草层的间接竞争力,可限制大部分矮小和浅根系杂草获取资源的能力,依托化感物质的直接竞争可抑制敏感型杂草的入侵、发育和存活,故一般杂草难以入侵、存活和造成危害。但常被侵占能力和繁殖能力极强的暖季型恶性杂草丝茅(Imperatakoenigii)入侵[8],一旦入侵难以彻底剔除,常在草坪中形成优势斑块或完全取代草坪草,造成草坪群落组成结构失衡、景观功能丧失和寿命缩短[9]。目前尚无有效、持久防除丝茅入侵和危害的技术措施。冷季型草坪草受生长方式、株丛类型和夏眠特性的限制,夏季生长停止、代谢缓慢、抗性降低,地上和地下竞争力衰退[10-11],极易因丝茅危害而退化。细叶结缕草和丝茅同为暖季型植物,夏季均为生长旺盛期,均有较强的拓展或侵占能力,丝茅如何入侵与危害结缕草草坪的机理尚不清楚。因此开展两者的竞争力及影响竞争力因素的研究,对该区细叶结缕草草坪建植和养护,防止丝茅入侵和制定合理防治方案具有一定的指导意义。
本试验依据暖季型草坪适宜温度为25~35 ℃,模拟该区夏季3个高温条件,对采用取代试验设计的5个不同丝茅侵入量的细叶结缕草草坪,进行30 d的间歇性高温冲击,之后通过测定其形态指标、生理指标和生物量,分析高温冲击对不同丝茅侵入量的结缕草草坪生长及竞争力影响。拟回答以下科学问题:1)高温冲击对两种植物地上、地下竞争力是否产生影响?2)哪个温度下丝茅攻击力最强,更利于其入侵和竞争?3)高温和侵入量对两种植物竞争力影响的差异?是否具有协同作用?4)高温冲击对两种植物生长、生理、再生的影响如何转变为潜在竞争力?5)两种植物现实竞争力和潜在竞争力的关系如何?旨在探讨抑制丝茅危害、提高草坪品质的有效途径,为草坪建植、养护、管理提供依据。
1 材料与方法
1.1 试验材料及设计
以细叶结缕草(Z.tenuifoliacv.Taiwan 2)及野生丝茅草为材料。采用高40 cm、口径30 cm塑料花盆,紫色土∶腐殖土(2∶1)为基质。
采用复合取代试验设计,于2017年7月,结缕草均匀播种、定苗100株/盆,待2~3真叶时,用丝茅草根茎插穗[长(2.0±0.2 cm),含1节、节左右各约1 cm、重约(0.15±0.01) g]替代部分幼苗,分别用丝茅每盆10、20、30、40、50株替代结缕草,形成结缕草∶丝茅草为9∶1,8∶2,7∶3,6∶4,5∶5的5个混播梯度,单播结缕草和丝茅草为对照(CK),每梯度种9盆,每梯度3盆为1组,分为3组。于(28±5) ℃的室内培养40 d后,于41~70 d,每天13:00-15:00,固定把每梯度3组分别移入温度为40、42、44 ℃的人工气候培养箱(光照强度1300 lx,相对湿度32%),进行高温冲击2 h,于71 d时进行测定。
1.2 测定项目与方法
构件性状:于71 d,每处理随机选结缕草、丝茅各10株,测定单株自然高度、分蘖数、叶片数、叶片厚度,用托普YMJ-C型叶面积仪测定总叶面积。
叶绿素含量:每盆随机剪取结缕草、丝茅草成熟健康叶片各0.2 g,采用乙醇丙酮混合提取法[12],测定叶绿素a和叶绿素b含量。
生理活性指标:每盆随机剪取结缕草、丝茅草成熟健康叶片各0.2 g,采用氮蓝四唑法[12]测定超氧化物歧化酶(superoxide dismutase, SOD)活性,采用愈创木酚法[12]测定过氧化物酶(peroxisome, POD)活性,采用紫外吸收法[13]测定过氧化氢酶(catalase, CAT)活性,采用硫代巴比妥酸法[13]测定丙二醛(malondialdehyde, MDA)含量,采用酸性茚三酮法[13]测定脯氨酸(proline, Pro)含量。
生物量:倾盆,分离,清洗植株,混播处理每盆随机选结缕草、丝茅各20株,单播对照组随机取40株,分离地上与地下部分,分别装袋,105 ℃下烘至恒重后称重,计算单株与每盆的地上、地下生物量。
相对产量:按照Fowler[14]的公式RYij=Yij/(pYi)和RYji=Yji/(qYj)计算。RYij代表与种j混播时种i的相对产量,RYji代表混播时种j的相对产量。Yij是与种j混播时种i的生物量,Yji是与种i混播播时种j的生物量,Yi是单播i的生物量,Yj是单播j的生物量。生物量均以盆为单位,而相对产量则以株为单位。p是混播方式下种i的比例,q为j的比例。当RY<1.0时,表示种间竞争大于种内竞争;当RY>1.0时,种内竞争大于种间竞争;而当RY=1.0时,表明种内和种间竞争水平相当。
相对产量总和:按公式RYT=pRYij+qRYji进行计算。RYT<1.0表明两物种间有拮抗作用,RYT>1.0表明两物种之间没有竞争,RYT=1.0表明两物种需要相同的资源,且一种可通过竞争将另一种排除出去。
攻击力指数:根据Mc Gilchrist等[15]的公式Ai=RYij-RYji和Aj=RYji-RYij计算。
竞争平衡指数:因两物种竞争平衡指数具有CBi=-CBj的关系[16],故按公式CBi=lnRYi/RYj计算种i的竞争平衡指数。CBi>0,说明物种i的竞争能力比物种j强;CBi=0,说明物种i和物种j竞争能力相等;CBi<0,说明物种i的竞争能力比物种j弱;CBi越大,说明物种i的竞争能力越强。
1.3 数据分析
用SPSS 19.0进行多重比较、方差分析等数据统计,并用Duncan法对各参数进行显著性检验。
选择18名空军工程大学在读大学生为被试,所有被试均为男性,年龄18~22岁(SD=3.08),被试均为右利手,视力或矫正视力正常,无色盲、色弱等特征,所有被试均要求有一定的电脑使用经验.
2 结果与分析
2.1 高温冲击对丝茅和结缕草现实竞争力的影响
2.1.1幼苗相对产量 高温冲击对丝茅和结缕草的地上和地下相对产量均产生显著影响(表1) (P<0.05)。随冲击温度升高,结缕草地上相对产量逐步下降(P<0.05),其地下相对产量先显著增加后急剧下降,地上和地下相对产量表现出不同消减趋势。丝茅的地上和地下相对产量则同步变化,42 ℃时丝茅地上和地下相对产量显著降低,44 ℃时地上和地下均达最大。随冲击温度升高,地上相对产量总和逐步下降(P<0.05),两种植物地上的拮抗作用越来越强,地下相对产量总和仅在44 ℃冲击下显著下降。
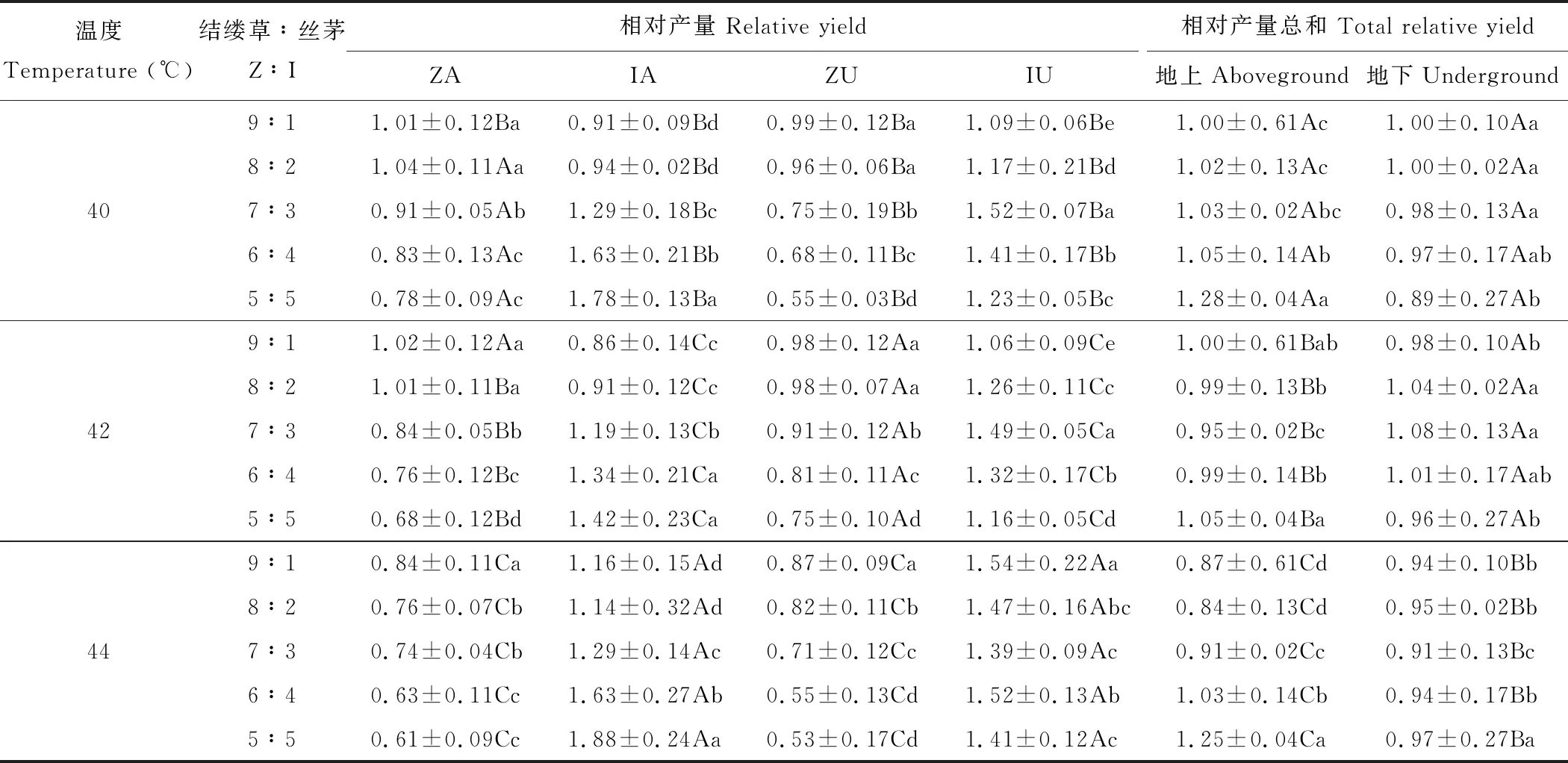
表1 高温冲击对丝茅和结缕草相对产量的影响Table 1 Effects of high temperature impact on relative competitiveness of I. koenigii and Z. tenuifolia
Z:Z.tenuifolia; I:I.koenigii; A: 地上生物量Aboveground biomass; U: 地下生物量Underground biomass. 同列不同大写字母表示冲击温度间差异显著(P<0.05),同列同温度的不同小写字母表示侵入量间差异显著(P<0.05)。 Different capital letters in the same column indicate significant differences in different temperature, different lowercase letters in the same column indicate significant difference in different proportion (P<0.05);下同The same below.
随丝茅侵入量增加,结缕草地上和地下相对产量不断下降(P<0.05),而在40和42 ℃时,≤20%丝茅入侵可显著促进结缕草地上生长(地上相对产量>1.00,结缕草种内竞争为主),对地下生长影响较小(地下相对产量近于1.00,种内与种间竞争近相等),此时丝茅地上产量受到抑制,丝茅地下产量则被促进。侵入量≥30%显著降低了结缕草地上和地下相对产量,且随冲击温度升高结缕草地上和地下相对产量下降越严重,种间竞争超过种内竞争。丝茅地上产量随侵入量增加显著增大,丝茅种内竞争加强,44 ℃侵入量50%时丝茅地上产量达1.88。丝茅地下产量在40和42 ℃时随侵入量增加表现为先增后减,30%侵入量丝茅地上产量高于其他侵入量。丝茅地下产量在44 ℃下侵入量10%时达最大值。
丝茅侵入量对地上和地下相对产量总和产生显著影响(P<0.05),在40 ℃时随侵入量增加地上相对产量总和增大且均大于1.00,地下相对产量总和则降低,≤30%侵入量地上和地下相对产量总和等于或近于1.00,此时两种植物需要相同的资源,可通过竞争将另一种排除,≥40%侵入量才显著增加地上相对产量总和,50%侵入量显著降低地下相对产量总和,表明两物种茎叶间无竞争,根系间则有拮抗作用。在42 ℃时30%侵入量的地上相对产量总和最小,20%~30%侵入量的地下相对产量总和则显著大于1.00。在44 ℃时40%和50%侵入量地上相对产量总和大于1.00,地下相对产量总和则均小于1.00。
方差分析表明,丝茅和结缕草的相对产量和相对产量总和,在总处理、侵入量间和冲击温度间均存在极显著差异(表2) (P<0.01)。结缕草地下相对产量比地上更易受生境影响,丝茅则相反。侵入量对结缕草和丝茅地上和地下相对产量的影响高于高温冲击。高温冲击和侵入量的影响大小均为结缕草地下产量>丝茅地上产量>结缕草地上产量>丝茅地下产量,互作对结缕草地下产量和丝茅地上产量具有极显著协同作用,对结缕草地上产量和丝茅地下产量具有显著作用。总处理、侵入量间、冲击温度间和互作间对地上相对产量总和影响均大于地下相对产量总和。
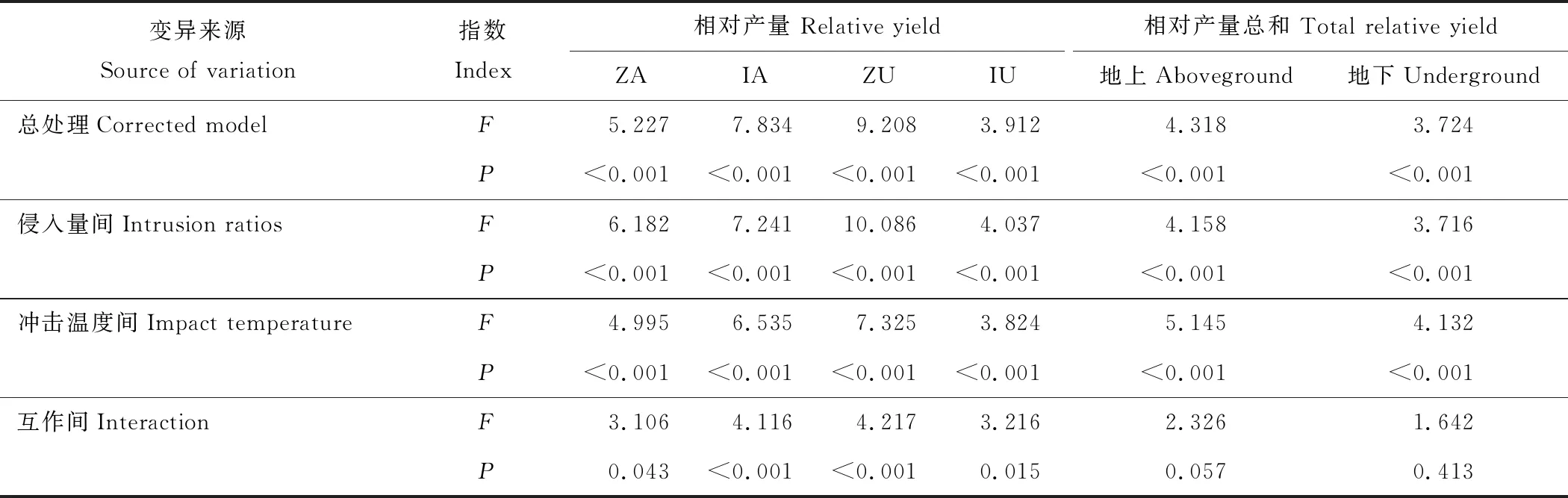
表2 丝茅和结缕草相对产量的方差分析Table 2 Variance analysis of the relative competitiveness of I. koenigii and Z. tenuifolia
注:F值表示F检验的显著性,F越大表示越显著,P值表示概率值,下同。
Note:Fvalue indicate the significance of theFtest, with greaterFvalue means the more significant, andPvalue indicates the probability value. The same below.
2.1.2攻击力和竞争平衡指数 高温冲击对结缕草地上和地下攻击力指数,及地上和地下竞争平衡指数产生显著影响(表3)(P<0.05)。随冲击温度升高,结缕草地上、地下攻击力指数和竞争平衡指数均为同步的先增后降趋势,即丝茅地上、地下攻击力指数和竞争平衡指数同步的先降后增,表明42 ℃时结缕草生长最旺盛、攻击力或抵抗丝茅攻击的能力最强。仅在40和42 ℃时侵入量≤20%条件下,结缕草地上攻击力指数和竞争平衡指数为正值,其余处理条件下结缕草攻击力指数和竞争平衡指数均为负值,可见丝茅对结缕草影响显著。
丝茅侵入量对结缕草地上、地下攻击力指数和竞争平衡指数均产生显著影响,随侵入量增加地上攻击力指数和竞争平衡指数显著下降,地下攻击力指数和竞争平衡指数则在侵入量50%时有所增加,40和42 ℃时侵入量30%条件下地下攻击力最小,44 ℃时侵入量40%下地下攻击力最小,表明丝茅对结缕草根系最大攻击力受温度和侵入量的协同影响。方差分析表明,总处理、侵入量间、冲击温度间和互作间,结缕草地上、地下攻击力指数和竞争平衡指数均存在极显著差异(表4)(P<0.01),地上部分比地下部分更易受侵入量和高温冲击的影响,侵入量对攻击力指数和竞争平衡指数的影响大于高温冲击。高温冲击对结缕草影响为地上攻击力指数>地上竞争平衡指数>地下竞争平衡指数>地下攻击力指数,侵入量影响为地上攻击力指数>地上竞争平衡指数>地下攻击力指数>地下竞争平衡指数,总处理影响为地上攻击力指数>地下竞争平衡指数>地上竞争平衡指数>地下攻击力指数。

表3 高温冲击对丝茅入侵的结缕草攻击力指数和竞争平衡指数的影响Table 3 Effects of high temperature impact on attack power and competition balance index of Z. tenuifolia invaded by I. koenigii

表4 结缕草攻击力指数和竞争平衡指数的方差分析Table 4 Variance analysis of the attack and competition balance index of Z. tenuifolia
2.2 高温冲击对丝茅和结缕草潜在竞争力的影响
2.2.1株高和分蘖数 高温冲击对丝茅和结缕草株高和分蘖数均产生显著影响(表5),丝茅的株高和分蘖数随冲击温度升高而显著增加,结缕草的株高和分蘖数在42 ℃后显著下降(P<0.05)。丝茅侵入量对丝茅和结缕草株高和分蘖数均有显著影响,随侵入量增加,丝茅株高显著提高而分蘖数降低,侵入量<30%对结缕草株高影响较小且一定程度上促进其分蘖,侵入量≥30%时显著增加结缕草株高而降低分蘖数。高温和侵入量总处理对株高和分蘖数具有极显著影响(P<0.01)(表6),方差分析结果表明,两种草的株高和分蘖数在冲击温度间、侵入量间、互作间均存在极显著差异(P<0.01),F值表明,侵入比例和互作对丝茅株高和分蘖数的影响大于结缕草,温度对结缕草株高和分蘖数的影响大于丝茅。

表5 高温冲击对丝茅和结缕草幼苗性状的影响Table 5 Effects of high temperature impact on seedling characteristics of I. koenigii and Z. tenuifolia
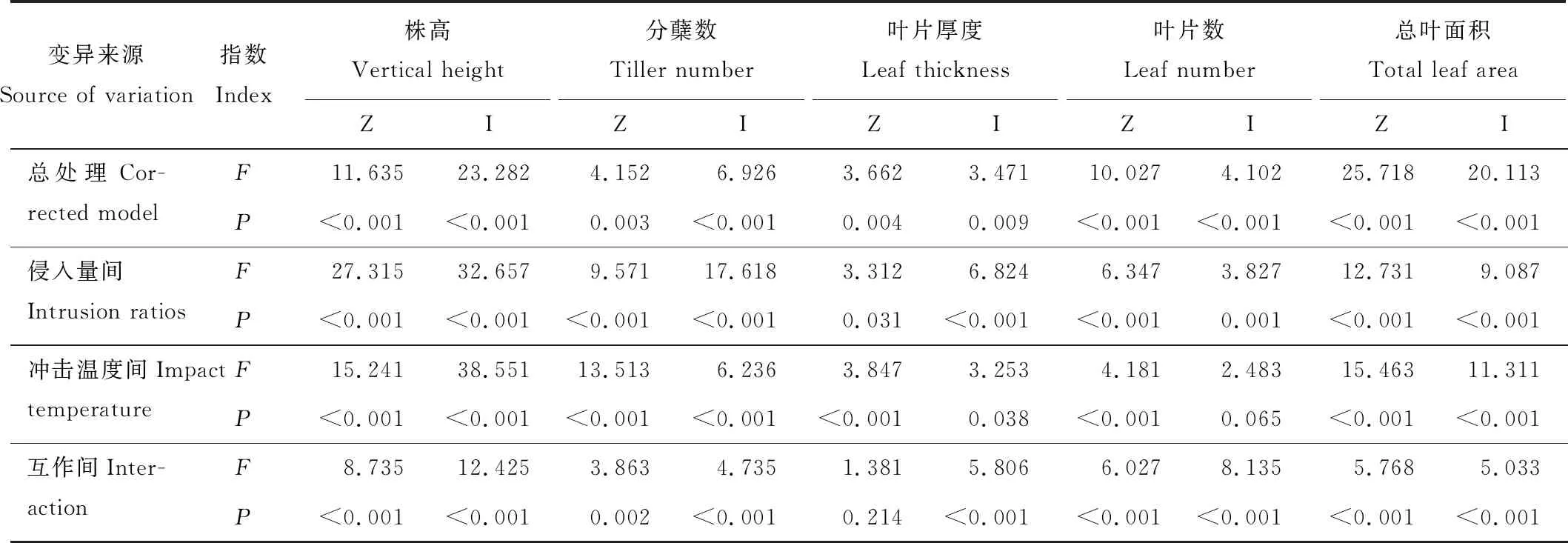
表6 丝茅和结缕草幼苗性状的方差分析Table 6 Variance analysis of the seedling characters of I. koenigii and Z. tenuifolia
2.2.3叶绿素含量 高温冲击对丝茅和结缕草的叶绿素含量均产生显著影响(表7)。随冲击温度升高,两者叶片中的叶绿素a和丝茅中叶绿素b含量显著增加(P<0.05)。38~42 ℃对结缕草叶绿素b含量影响较小,而44 ℃下显著提高了叶绿素b含量。丝茅中叶绿素a/b随温度增加而显著降低(P<0.05),42 ℃结缕草叶绿素a/b达最大值,40和44 ℃间差异较小。3个冲击温度下,丝茅侵入量≥20%均显著降低了结缕草叶绿素a和叶绿素b含量,而叶绿素a/b随侵入量增加而增大。≤20%侵入量显著提高了丝茅叶绿素含量,>20%后丝茅叶绿素含量随冲击温度不同表现为增加或减少。

表7 高温冲击对丝茅和结缕草叶绿素含量的影响Table 7 Effects of high temperature impact on chlorophyll content of I. koenigii and Z. tenuifolia
方差分析表明,丝茅和结缕草叶绿素含量在总处理、侵入量间和冲击温度间均存在极显著差异(表8) (P<0.01)。总处理对两种植物的影响叶绿素a>叶绿素b>叶绿素a/b,对结缕草叶绿素a和叶绿素a/b的影响强于丝茅。温度对丝茅叶绿素含量影响大于结缕草,侵入量对结缕草的叶绿素a和丝茅的叶绿素b含量影响较大。结缕草叶绿素受影响顺序为侵入量间>冲击温度间>互作间,丝茅叶绿素a主要受温度影响,叶绿素b主要受比例影响。
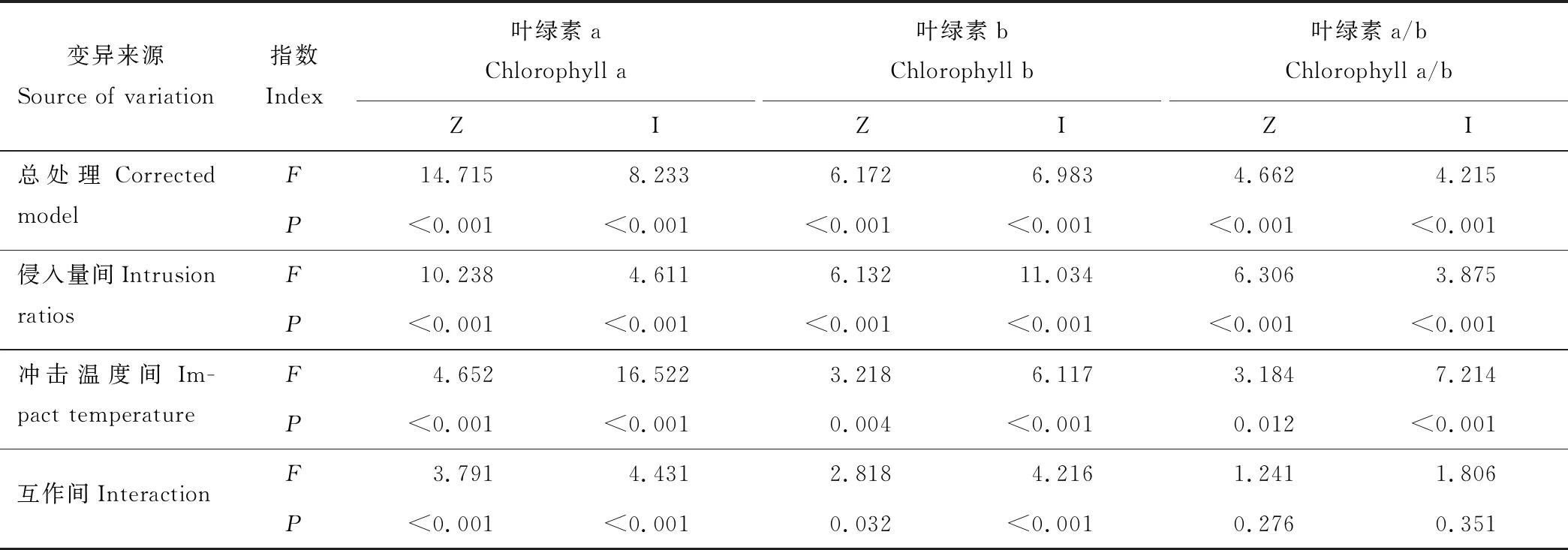
表8 丝茅和结缕草叶绿素含量的方差分析Table 8 Variance analysis of the chlorophyll content of I. koenigii and Z. tenuifolia
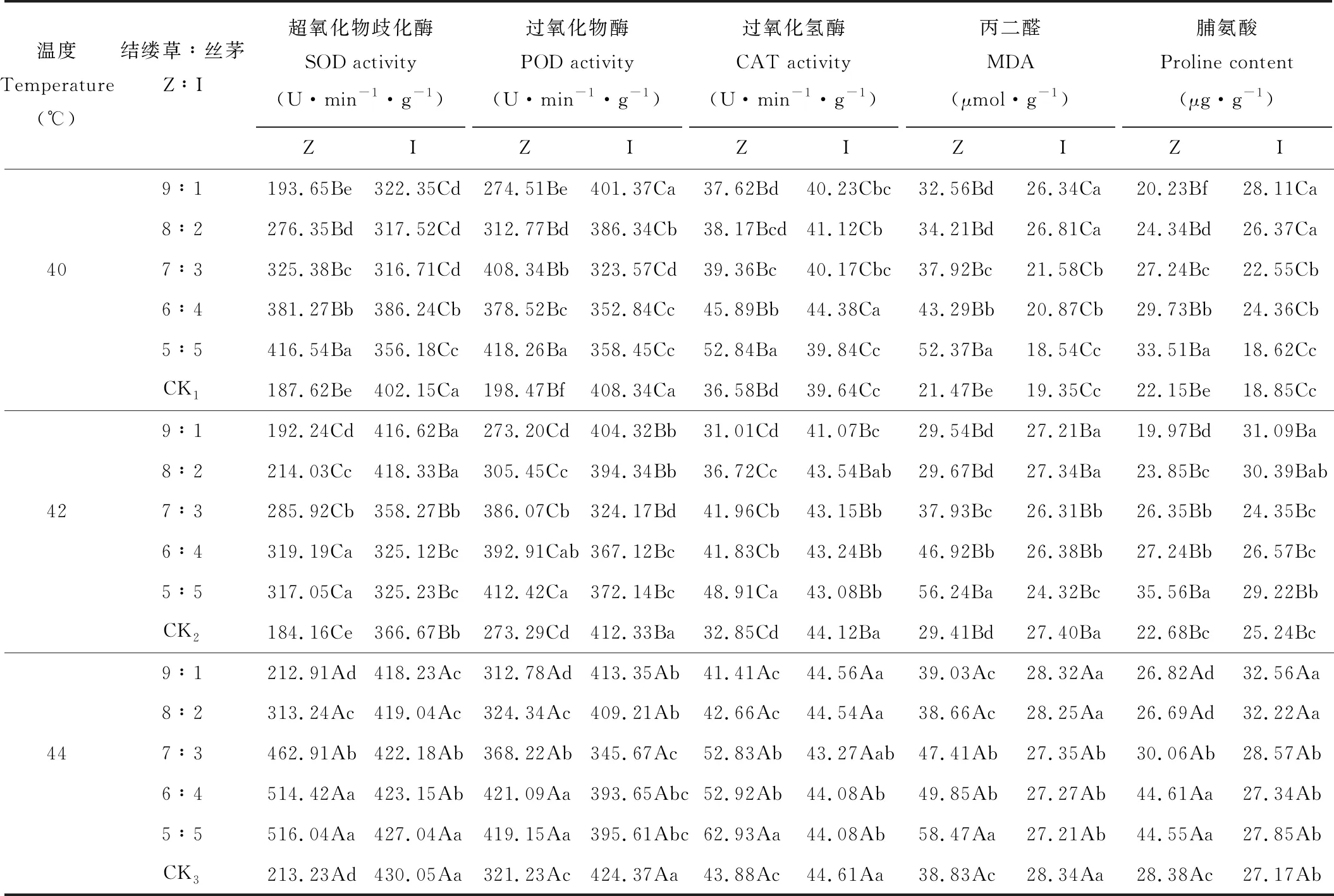
表9 高温冲击对丝茅和结缕草抗性物质的影响Table 9 Effects of high temperature impact on resistant substance of I. koenigii and Z. tenuifolia
2.2.4抗性物质 高温冲击对丝茅和结缕草的抗性物质含量均产生显著影响(表9)。随冲击温度升高,丝茅的SOD、POD、CAT活性和MDA、Pro含量均显著增加(P<0.05)。结缕草的SOD、POD、CAT活性在42 ℃时显著低于其他温度,44 ℃时显著高于其他温度,MDA、Pro含量在40与42 ℃间差异较小,在44 ℃时急剧增加。
3个冲击温度下,丝茅侵入量≥20%时均显著增加了结缕草SOD、POD、CAT活性(P<0.05)。40 ℃冲击下,侵入量≥10%时显著增加了结缕草MDA、Pro含量,42和44 ℃冲击下,侵入量≥30%, MDA、Pro含量增加。且10%侵入量下,3个高温冲击显著降低了结缕草Pro含量(P<0.05)。丝茅侵入量对丝茅抗性物质含量的影响,随冲击温度不同而不同,40 ℃下不同侵入比例的丝茅SOD和POD活性低于对照,CAT活性和MDA、Pro含量则>对照。42和44 ℃冲击下,丝茅POD、CAT活性和MDA含量均低于对照,Pro含量在侵入量≤20%时<对照。
方差分析表明,丝茅和结缕草的抗性物质,在总处理、侵入量间和冲击温度间均存在极显著差异(表10)(P<0.01)。由F值可见,温度对结缕草抗性物质影响大于丝茅,侵入比例对结缕草SOD、POD、CAT活性和MDA含量影响大于丝茅,对Pro含量则小于丝茅。结缕草抗性物质受温度影响大于侵入量,温度对结缕草抗性物质的影响为SOD>MDA>CAT>Pro>POD,侵入量的影响为SOD>MDA>CAT>POD>Pro。温度对丝茅影响为SOD>POD>MDA>CAT>Pro,侵入量的影响为SOD>Pro>POD>MDA>CAT。

表10 丝茅和结缕草抗性物质含量的方差分析Table 10 Variance analysis of the resistant substance of I. koenigii and Z. tenuifolia
3 讨论
研究混生种群的竞争行为和物种间相互作用的试验设计,有部分增加、取代系列、添加系列、完整添加等方式[17],每种设计都有一定的优缺点。取代系列设计在研究两物种竞争能力、相互作用类型、生态位分化和环境资源利用效率方面具有优势[18],但仅以地上生物量为基础进行两物种的地上部分竞争力评价,不能反映地下部分的竞争强度。考虑到草坪结构、功能及养护措施的特殊性,地下部分对养分吸收和分生再生能力的重要性,故本试验在采用种群密度不变、只改变种群组成的复合取代设计基础上,研究草坪植物竞争时,同时测定地上、地下生物量分析两物种的立体竞争[19]。高温冲击对两种植物地上、地下竞争力均有显著影响,加剧了两种植物地上部分的拮抗作用。随冲击高温升高结缕草的地下相对产量不断下降,丝茅的地下相对产量不断增加。随混生群落中丝茅比例增大,结缕草地下相对产量进一步降低。根系吸收养分能力降低而地上相对产量增加的失衡生长,使结缕草在高温与丝茅的双重胁迫下,攻击力和竞争平衡参数急剧下降,面临被排挤出群落的风险。
竞争力受物种生物学特性影响,更受生长发育阶段及环境因子制约[20]。细叶结缕草匍匐生长、70%的纤细根系分布在20 cm以内,丝茅兼有匍匐和直立生长、粗壮根系可深达2 m左右,丝茅的抗逆性结构基础优于结缕草。本研究以相对产量表征两种植物竞争力,而丝茅的单株的地上、地下部分绝对产量大于结缕草,随冲击高温增加和丝茅侵入量增大,结缕草相对产量降低的同时,失衡生长使地下部分绝对产量降低,根系分生和更新能力下降直接导致其竞争力衰退。随幼苗成长和有性繁殖形成,丝茅以化感作用表达的直接竞争不断增强[21],结缕草将面临更大的竞争压力。丝茅对结缕草攻击力受生境温度和侵入量的影响,在室温下,侵入量≥20%时,丝茅种内竞争大于种间竞争,对结缕草地上相对产量、地上攻击力指数和竞争平衡指数几无影响[22]。≥40 ℃的高温使结缕草生长受阻,混生群落的种间竞争大于种内竞争,侵入量代替高温冲击成为影响结缕草竞争力的主要原因。
取代设计基于两种植物现有生物量进行现实竞争力评价,未考虑生物量结构与构件分配比,是过去两种植物资源侵占与能量累积的结果。相同现有生物量,若根、茎、叶生物量分配比例不同,引起叶面积、株高、分蘖数、茎基和根系差异,影响植株的光合同化、空间拓展、分生再生和抗逆能力,决定植株现实生长情况及将来的竞争趋向。高温冲击增加了丝茅的株高和分蘖数、叶片数和叶面积,随冲击温度升高,丝茅的空间优势、光合同化优势和再生能力不断提升。高温冲击限制了结缕草生长,丝茅入侵量抑制了结缕草叶面积、分蘖数和株高,使结缕草植株低矮、纤细、叶少而薄,对光的捕获与同化受限。高温冲击和丝茅入侵降低了结缕草的叶绿素含量,叶绿素含量与植株光合能力及抗逆性密切相关,叶绿素含量下降是叶片衰老和光合速率下降最明显的特征[23]。高温冲击提高了丝茅和结缕草的抗氧化酶活性,SOD首先将多余的超氧阴离子结合成H2O2[24],POD和CAT则进一步分解为无害的H2O和O2[25],结缕草抗氧化酶活性受冲击温度影响大于丝茅,丝茅入侵进一步使结缕草将更多的能量用于抵御胁迫。高温冲击和丝茅入侵提高了结缕草细胞膜透性,随Pro和MDA含量增加结缕草细胞受伤害程度加剧。丝茅的Pro和MDA含量受冲击温度影响,更受丝茅侵入量影响,或以密度为基础的种内竞争是决定丝茅细胞膜透性的主要因子,需深入研究。
高温冲击对受丝茅入侵的细叶结缕草现实和潜在竞争力的影响,使结缕草植株矮化、枝条密度稀疏化、根系浅表化,分生再生能力降低甚至丧失。丝茅强大的平衡生长能力、生理整合能力和繁殖扩散能力,在时间、空间与环境因子的契合作用下,其潜在优势必将转化为现实的攻击力和竞争优势,对结缕草产生持续的竞争效应。丝茅根茎含有醇、萜醇、甾醇、酸类等[26]次生代谢产物,对坪床土壤的含水量、容重、pH均有显著的影响[27],大量与长期入侵必将对结缕草根系产生化感作用或拮抗作用,使根系分生、更新、吸收能力下降,导致结缕草适合度、优势度、存活率与繁殖率等特征指数变化,最终草坪的组成、结构与功能发生改变。
4 结论
1)高温冲击加剧了两种植物地上部分的拮抗作用,引起了结缕草地上与地下生长失衡,≥40 ℃温度显著降低结缕草地下竞争力。仅40和42 ℃下、侵入量≤20%时,结缕草地上竞争力占优势,其他温度和侵入量下丝茅地上和地下攻击力大于结缕草。
2)高温冲击降低了结缕草叶片数和叶面积,>42 ℃高温限制了结缕草分蘖数,≥30%丝茅侵入量抑制结缕草分蘖、降低了结缕草叶片厚度、叶片数和叶面积,高温冲击和丝茅入侵对结缕草叶绿素含量、SOD、POD、CAT活性和MDA、Pro含量有显著影响,使结缕草光合同化、空间拓展、分生再生和抗逆能力等潜在竞争力下降。
3)高温冲击对结缕草形态、生理、相对产量及攻击力的影响大于丝茅,加剧了丝茅入侵对结缕草现实竞争力的影响,高温和丝茅侵入量协同作用限制了结缕草潜在竞争力,潜在竞争力和现实竞争力互为因果相互转化,故控制丝茅侵入量≤20%、降低生境温度是夏季结缕草草坪养护的基本要求。
猜你喜欢
今日农业(2022年16期)2022-11-09 23:18:44
今日农业(2022年15期)2022-09-20 06:55:48
环球时报(2022-06-20)2022-06-20 17:06:23
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
中国果业信息(2019年1期)2019-01-05 17:41:42
基层中医药(2018年8期)2018-11-10 05:32:06
生物学教学(2017年9期)2017-08-20 13:22:32
汽车维修与保养(2015年12期)2015-04-18 07:51:45
汽车维修与保养(2015年6期)2015-04-17 03:31:44
汽车维护与修理(2015年6期)2015-02-28 12:17:08
