
苗期低温胁迫下烯效唑对红小豆根系抗寒生理及产量的影响
2019-07-27 01:44:36项洪涛李琬何宁王雪扬郑殿峰王彤彤梁晓艳唐晓东李一丹
草业学报 2019年7期
项洪涛,李琬,何宁*,王雪扬,郑殿峰,王彤彤,梁晓艳,唐晓东,李一丹
(1.黑龙江省农业科学院耕作栽培研究所,黑龙江 哈尔滨 150086;2.黑龙江八一农垦大学农学院,黑龙江 大庆163319)
红小豆(Vignaangularis),又名红豆、赤小豆、小豆等,是豆科豇豆属一年生草本短日照植物[1],富含蛋白质、铁、维生素等营养成分,属于药食兼用作物。红小豆对环境适应性很强,在瘠薄、盐碱和干旱土壤上均可生长,但其不耐水涝和冷害[2]。红小豆对温度较为敏感、喜光而不耐冷[3],最适生育温度是20~24 ℃。温度是影响作物生长发育的重要气象因素之一[4],生育期遭受低温会引起植物体内生理代谢发生明显变化[5]。在全球气候变暖的背景下,极端天气事件出现的频率呈现增多增强的趋势[6],近年来我国东北地区春季和夏季频繁发生的低温,对农业生产造成很大影响。Badowiec等[7]指出,在影响作物生产力的诸多因素中,冷害是其中重要一项,其干扰作物的生理生化过程,制约作物产量和品质[8]。相同作物不同品种之间对低温的应答存在很大区别,不同低温持续时间对同一品种的影响也完全不同[9]。冷害导致豆类作物减产有3个主要原因,一是苗期冷害延缓前期的生长发育;二是花期冷害影响叶片光合产物的输出,致使花和荚大量脱落;三是成熟期冷害造成籽粒充实不良[10]。郭数进等[10]表明苗期冷害导致豆类作物减产的原因主要是延迟了植株前期的生长发育。王萍等[11]指出开花期遭遇冷害会导致豆科作物荚数减少,籽粒空瘪,最终减少产量。张勇[12]研究表明冷害使单株粒数及百粒重降低明显,致使产量明显降低,减产幅度与受冷害程度呈正相关。
烯效唑(uniconazole),简称S3307,是一种经实践证明相当高效的植物生长延缓剂[13],具有广谱、高效、低毒、低残留的特点。S3307有提高产量和品质的作用,常被应用在调控作物生长和发育、增强抗逆性等方面。有报道指出,S3307具有缓解逆境胁迫的效果并显著提高产量[14];另有研究表明,低温条件下,烯效唑灌根处理能提高苗木质膜相对透性,有助于提高苗木的抗寒能力[15];曲亚英等[16]指出,低温条件下,S3307浸种显著增强甜椒幼苗的可溶性物质和脯氨酸含量,缓解低温伤害;郑春芳等[17]指出,S3307能显著提高低温胁迫下超氧化物岐化酶(superoxide,SOD)和过氧化物酶(peroxidase,POD)活性,增强秋茄的抗逆能力,从而提高秋茄幼苗的抗冷性。这些研究说明S3307具有提高植物抗冷的效应,但目前对缓解豆科作物低温伤害的研究很少,尤其是低温影响红小豆苗期根系生理机能的变化及S3307提高其抗冷的研究未见报道。因此,本试验于幼苗期对红小豆进行低温处理,开展低温胁迫下S3307对红小豆根系生理及产量的影响,旨在分析S3307抵御低温的作用,丰富东北地区红小豆抗冷技术工程体系,为红小豆耐冷育种、高产优质生产提供理论依据和技术支持。
1 材料与方法
1.1 试验材料
选用宝清红(耐冷)和天津红(冷敏)两个温度敏感性存在显著差异的红小豆为试验品种。供试材料由国家食用豆产业技术体系栽培生理岗位实验室提供。
供试植物生长调节剂为S3307,由黑龙江八一农垦大学化控研究中心提供。
1.2 试验设计与处理
试验于2017和2018年在黑龙江省农业科学院耕作栽培研究所盆栽场及人工气候室内进行,由于两年规律一致,本研究选用2018年数据。采用盆栽方式,每盆保苗3株,试验用盆高30 cm、直径25 cm。试验用土取自哈尔滨市道外区民主乡,土壤类型为草甸黑土。供试土壤的理化性质:土壤容重约为1.67 g·cm-3,最大田间持水量约为26.88%,有机质含量2.92%、全氮1.35 mg·g-1、全磷0.64 mg·g-1、缓效钾341.5 mg·kg-1、碱解氮151.6 mg·kg-1、速效磷49.97 mg·kg-1、速效钾149.6 mg·kg-1、土壤pH 6.77。2018年4月30日播种,每品种分别播种150盆,选取长势均匀的样本进行试验,试验共设6个处理,每个处理设3次重复,具体设计见表1。待植株生长至幼苗期时(真叶完全展开期,5月13日)进行低温处理,处理温度为恒定15 ℃。开始低温处理当天上午10:00,叶面喷施20 mg·L-1S3307(每hm2用液量为225 L)。喷施完毕后随即进入人工气候室内进行低温处理,持续时间分别是1,2,3,4,5 d。
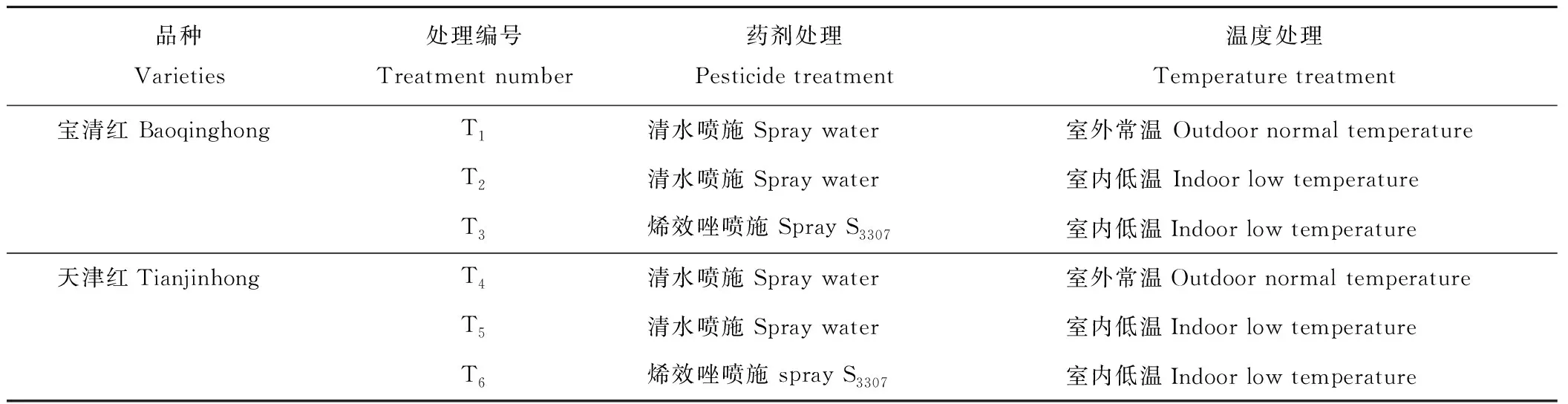
表1 试验设计Table 1 Experiment design
1.3 测定项目及方法
1.3.1取样方法 处理期间,连续取样5次,每天上午10:30取样一次,各处理分别进行取样,将根系样品充分清洗并吸干水分后进行取样,并立即放入液氮中,而后置于-80 ℃冰箱中保存,供测定生理指标使用。
低温处理后每天各处理移至室外6盆,直至成熟,供产量调查使用。
1.3.2测定方法 按照李合生等[18]的方法测定超氧化物歧化酶、过氧化物酶、过氧化氢酶(catalase,CAT)活性及丙二醛(malondialdehyde,MDA)含量;采用考马斯亮蓝G-250染色法测定可溶性蛋白含量[18];采用硫酸蒽酮比色法测定可溶性糖含量[19];采用茚三酮比色法测定脯氨酸含量[19];百粒重采用称重法;采用称重法以每盆为单元测定产量。
1.4 数据处理
采用Excel 2010进行数据处理和作图,使用DPS软件进行统计分析。
2 结果与分析
2.1 幼苗期低温胁迫下S3307对红小豆根系膜脂过氧化和渗透调节的影响
2.1.1对根系MDA含量的影响 幼苗期低温处理后,红小豆根系内MDA含量呈增加趋势,随低温处理时间的延长,MDA含量随之增高,冷敏品种天津红的MDA含量高于耐冷品种宝清红(图1)。经方差分析可知:宝清红低温处理1~3 d时,T2、T3与T1相互之间没有显著性差异;低温处理5 d时,T2较T1高45.41%,达显著差异水平。天津红低温处理1~2 d,T6与T5之间没有明显差异;处理3 d时,T5显著高于T6和T4,但T6和T4间无显著差异;处理4 d时,T5显著高于T6和T4,但T6和T4间无显著差异;处理第5天,T5较T4高51.08%,差异达显著水平。S3307能够明显抑制低温条件下MDA含量的增加,低温处理5 d时,T3较T2低9.05%、T6较T5低4.37%。

图1 幼苗期低温条件下S3307对红小豆根系丙二醛含量的影响Fig.1 Effect of S3307 on MDA content under low temperature stress in root of adzuki bean during seedling stage 采用新复极差法进行多重比较,不同字母表示同一天不同处理间在0.05水平差异显著,下同。Duncan’s multiple range test. Different letters indicate significant difference at 0.05 level under different treatments at the same day. The same below.
2.1.2对根系脯氨酸含量的影响 幼苗期低温导致红小豆根系中脯氨酸含量呈先升后降的变化趋势,整体看耐冷品种的脯氨酸含量高于冷敏品种(图2)。叶面喷施S3307能够在一定时间内促进脯氨酸含量增加,并抑制脯氨酸下降速率。低温处理导致红小豆根系脯氨酸含量在第3天上升到最高,而后开始降低,处理第3天时,T2较T1高10.12%、T5较T4高25.39%;处理第5天时,T2较T1低8.34%、T5较T4低4.90%,方差分析表明差异均未达到显著水平。但喷施S3307后,脯氨酸含量上升,到处理第4天后开始下降,低温处理5 d时,T3较T2高20.30%、T6较T5高出7.79%。说明喷施S3307能够促进脯氨酸含量的提高并推迟脯氨酸含量下降的时间。

图2 幼苗期低温条件下S3307对红小豆根系脯氨酸含量的影响Fig.2 Effect of S3307 on proline content under low temperature stress in root of adzuki bean during seedling stage
2.2 幼苗期低温胁迫下S3307对红小豆抗氧化酶系统的影响
2.2.1对根系SOD活性的影响 随着幼苗期低温处理时间的延长,红小豆根系SOD活性呈先升后降的变化规律(图3)。宝清红和天津红均于处理第3天达到最大值。处理第3天时,T2较T1高14.92%、T5较T4高11.01%;处理第5天时,T2较T1低15.27%、T5较T4低6.42%,方差分析表明差异均未达到显著水平。S3307能明显提高SOD活性,从低温处理第2天开始到第5天,T3处理持续高于T2,T6处理持续高于T5;其中处理第3天时,T3较T1和T2分别高出30.37%和13.45%、T6较T4和T5分别高出37.72%和24.06%;处理第5天时,T3较T2高出18.48%、T6较T5高出10.40%。整体分析,S3307对天津红的提高幅度更大,提高SOD活性的效应更为明显。
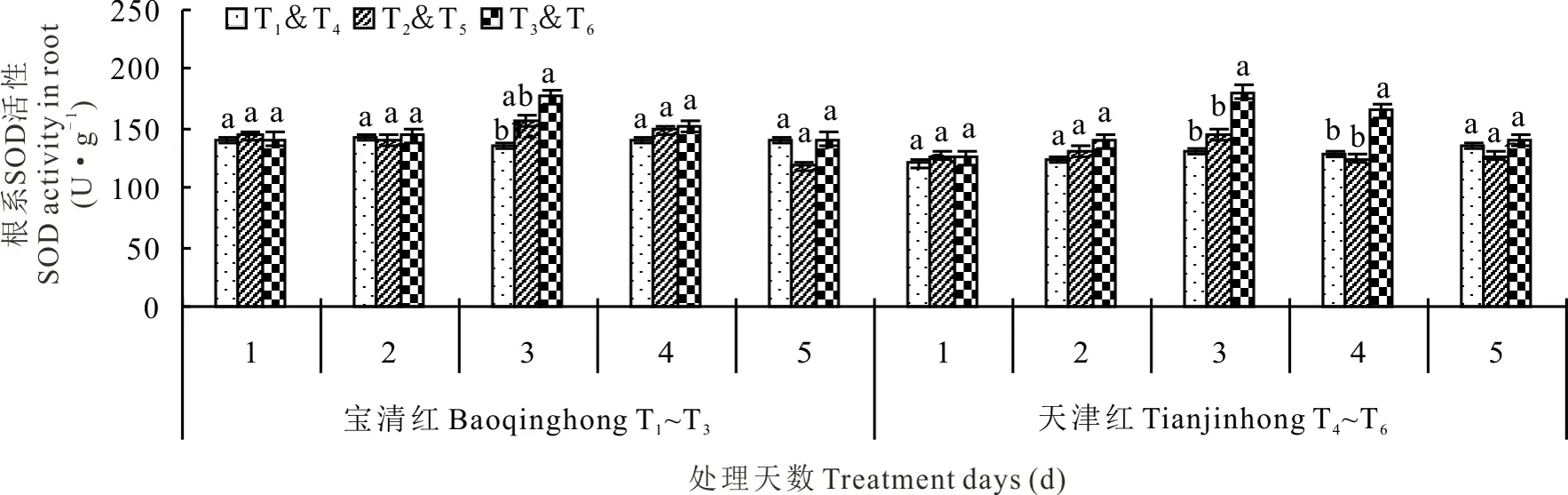
图3 幼苗期低温条件下S3307对红小豆根系SOD活性的影响Fig.3 Effect of S3307 on SOD activity under low temperature stress in root of adzuki bean during seedling stage
2.2.2对根系POD活性的影响 幼苗期低温处理后,红小豆根系POD活性先升高后降低,T2和T5都是在处理后第2天达到最高值(图4)。处理第2天时,T2较T1显著高出40.11%、T5较T4高8.25%;处理第3天时,T2较T1高5.93%,T5较T4高0.75%;处理第5天时,T2较T1低17.38%、T5较T4低14.03%,方差分析表明差异均未达到显著水平。而喷施S3307后,T3在第3天达到峰值,但T6仍是在第2天达到峰值,处理第2天时,T3较T2显著低14.51%、而T6较T5显著高23.48%;低温处理3 d,T3较T2显著高出23.60%、T6较T5显著高出15.95%;处理第5天,T3较T2显著高出35.30%、T6较T5高出17.72%。整体看喷施S3307可提高红小豆根系POD活性,1~3 d对冷敏品种天津红效果明显,而3~5 d对宝清红效果更明显。
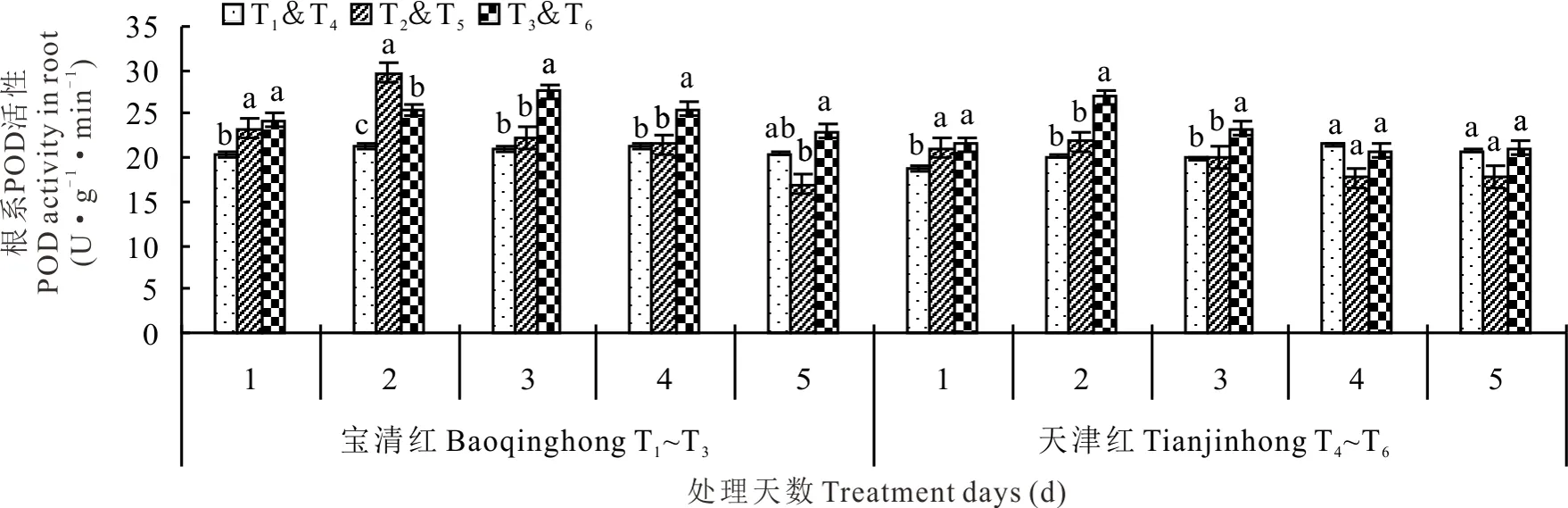
图4 幼苗期低温条件下S3307对红小豆根系POD活性的影响Fig.4 Effect of S3307 on POD activity under low temperature stress in root of adzuki bean during seedling stage
2.2.3对根系CAT活性的影响 幼苗期低温处理后,红小豆根系内CAT活性呈单峰曲线的变化趋势,宝清红的CAT活性峰值出现在低温处理后第3天,而天津红在低温处理后第4天达到最高值(图5)。处理第3天时,T2较T1高出53.33%、T5较T4高13.33%;处理第4天时,T2较T1高17.65%,T5较T4高6.25%;处理第5天时,T2较T1高6.67%、而T5较T4低12.50%,方差分析表明差异均未达到显著水平。喷施S3307能够明显促进CAT活性的提高,低温处理期间,T3持续高于T2,T6持续高于T5。处理第4天时,T3较T2高15.00%,T6较T5高17.65%;处理第5天时,T3较T2显著高出25.00%、T6较T5高28.57%。整体来看,低温处理后宝清红的CAT活性高于天津红,S3307对宝清红的调控效应更为明显。
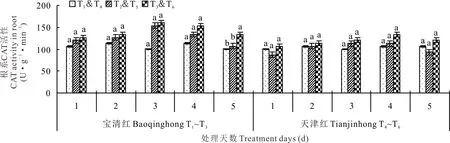
图5 幼苗期低温条件下S3307对红小豆根系CAT活性的影响Fig.5 Effect of S3307 on CAT activity under low temperature stress in root of adzuki bean during seedling stage
2.3 幼苗期低温胁迫下S3307对红小豆可溶性物质含量的影响
2.3.1对根系可溶性糖含量的影响 幼苗期低温处理后,红小豆根系可溶性糖含量呈先升后降的变化趋势,其中T2在低温第4天时达到峰值,T5在第3天达到峰值(图6)。处理第2天时,T2较T1低10.84%、T5较T4高2.85%;处理第3天时,T2较T1高18.97%,T5较T4高17.55%;处理第4天时,T2较T1显著高出86.14%,T5较T4显著低15.74%;处理第5天时,T2较T1显著低46.24%、而T5较T4显著低42.01%。喷施S3307可提高红小豆根系可溶性糖含量,尤其以处理后4~5 d更为明显。处理第3天时,T3较T2高1.48%,而T6较T5显著低了17.38%;处理第4天时,T3较T2显著高出17.34%,而T6较T5显著高出30.71%;处理第5天时,T3较T2显著高出120.95%、而T6较T5显著高出125.27%。
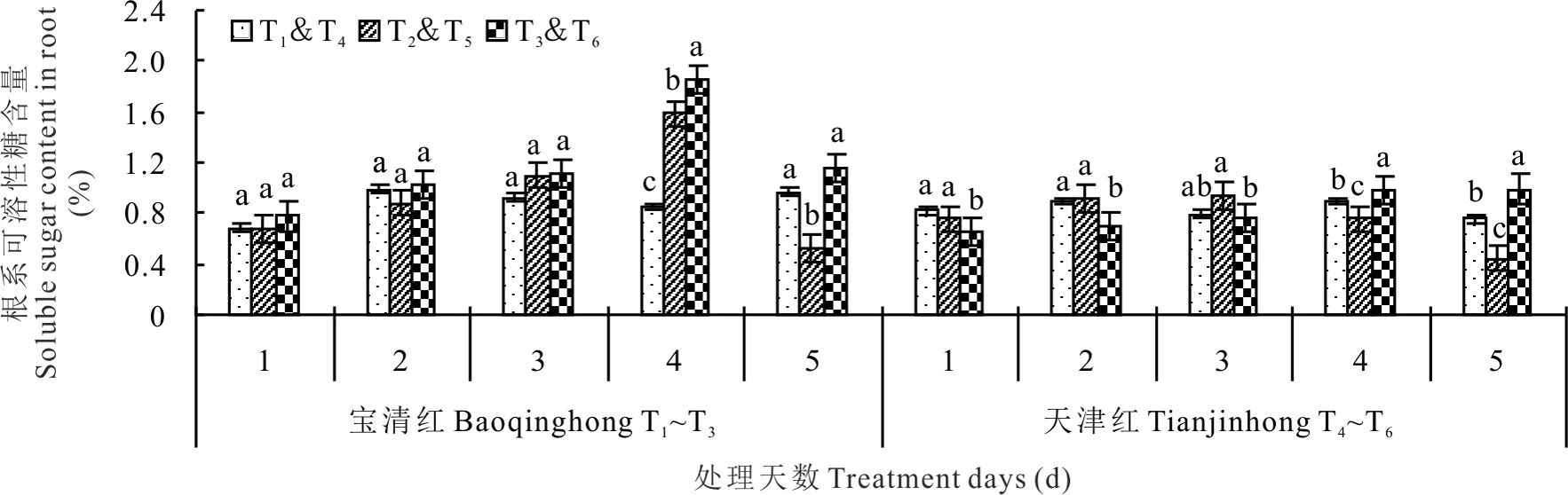
图6 幼苗期低温条件下S3307对红小豆根系可溶性糖含量的影响Fig.6 Effect of S3307 on soluble sugar content under low temperature stress in root of adzuki bean during seedling stage
2.3.2对根系可溶性蛋白含量的影响 由图7可知,幼苗期低温处理后,红小豆根系可溶性蛋白质含量呈单峰曲线变化规律,T2和T5均在低温第2天时达到最高值。处理第2天时,T2较T1显著高出5.30%,T5较T4显著高11.10%;处理第3天时,T2较T1显著高出6.27%、T5较T4显著高3.15%;处理第4天时,T2较T1显著低了6.92%,T5较T4显著低了3.15%;处理第5天时,T2较T1显著低21.67%、而T5较T4显著低35.63%。喷施S3307能够明显促进低温条件下红小豆根系可溶性蛋白质的提高,低温处理3 d时,T3较T2高出2.62%,T6较T5显著高出3.91%;处理第4天时,T3较T2显著高出8.89%,T6较T5显著高出22.56%;处理第5天时,T3较T2显著高出11.43%,T6较T5显著高出13.64%。低温条件下,喷施S3307可提高红小豆根系可溶性蛋白质含量,尤其是低温处理后第3~5天较为明显。
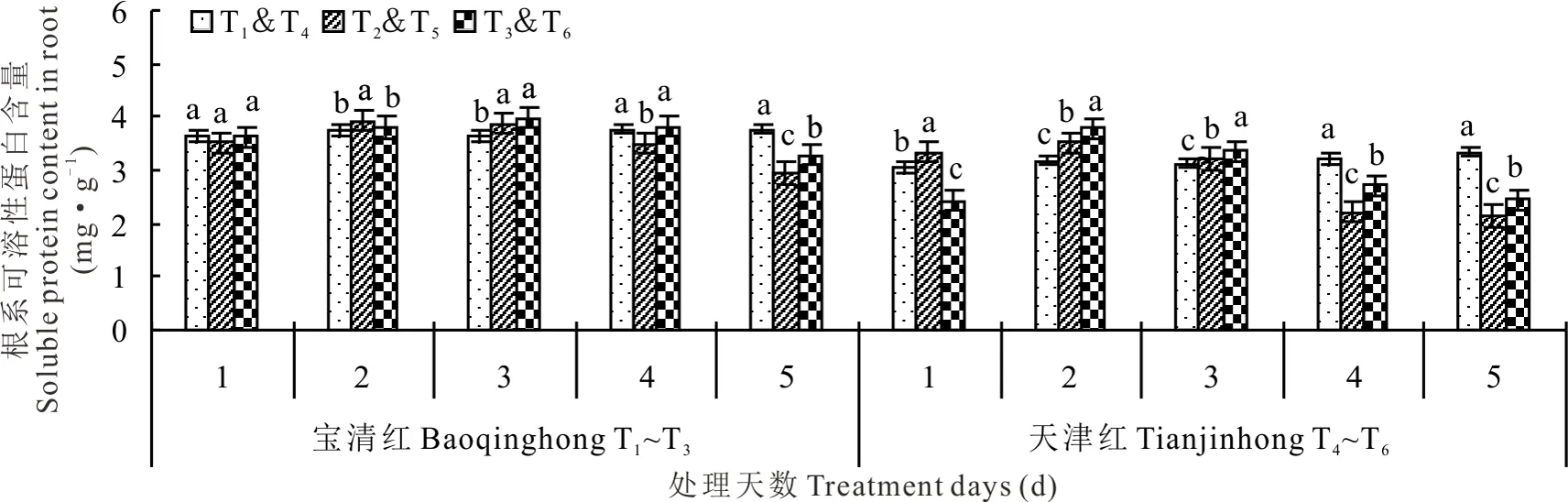
图7 幼苗期低温条件下S3307对红小豆根系可溶性蛋白质含量的影响Fig.7 Effect of S3307 on soluble protein content under low temperature stress in root of adzuki bean during seedling stage
2.4 幼苗期低温胁迫下S3307对红小豆产量的影响
2.4.1对红小豆百粒重的影响 幼苗期低温处理后,红小豆百粒重略有下降,T2重量低于T1,T5低于T4(表2)。低温条件下,喷施S3307可提高红小豆的百粒重,尤其是低温处理后第3~5天较为明显。方差分析可知:宝清红低温处理3 d时,T3显著高于T2;处理4和5 d时,T1、T2和T3之间差异不显著。天津红低温处理4 d时,T6显著高于T5;其他各处理时间,T4、T5和T6之间无显著性差异。
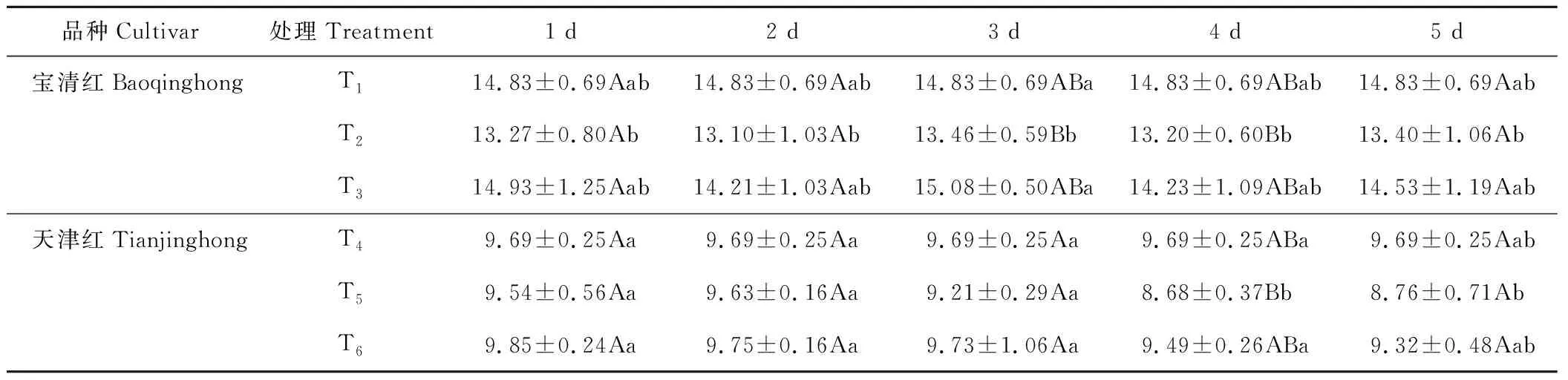
表2 幼苗期低温条件下S3307对红小豆百粒重的影响Table 2 Effect of S3307 on hundred-grain weight under low temperature stress of adzuki bean during seedling stage (g)
注:数据为平均值±标准差。采用新复极差法进行多重比较,同列小写字母表示在0.05水平显著,大写字母表示在0.01水平显著,下同。
Note: Date is mean±SD. Treatment means were separated using Duncan’s multiple range test. Small and capital letters within each column indicate difference at 0.05 and 0.01 levels of probability, respectively. The same below.
2.4.2对红小豆产量的影响 幼苗期低温处理后,红小豆产量降低(表3)。T2产量低于T1,T5低于T4。处理第5天时,T2较T1下降了19.39%、T5较T4下降了41.69%。方差分析结果表明,宝清红处理3 d时,T2显著低于T1;天津红处理2和3 d时,T5显著低于T4,处理4和5 d,T5极显著低于T4。
低温条件下,喷施S3307可提高红小豆产量,尤其是低温处理3 d后更加明显,低温处理第5天时,T3较T2提高了23.06%,T6较T5提高了43.88%。经方差分析可知:宝清红低温处理3 d时,T3显著高于T2,但未达到极显著差异水平。天津红低温处理4 d时,T6显著高于T5;低温处理5 d时,T6极显著高于T5。
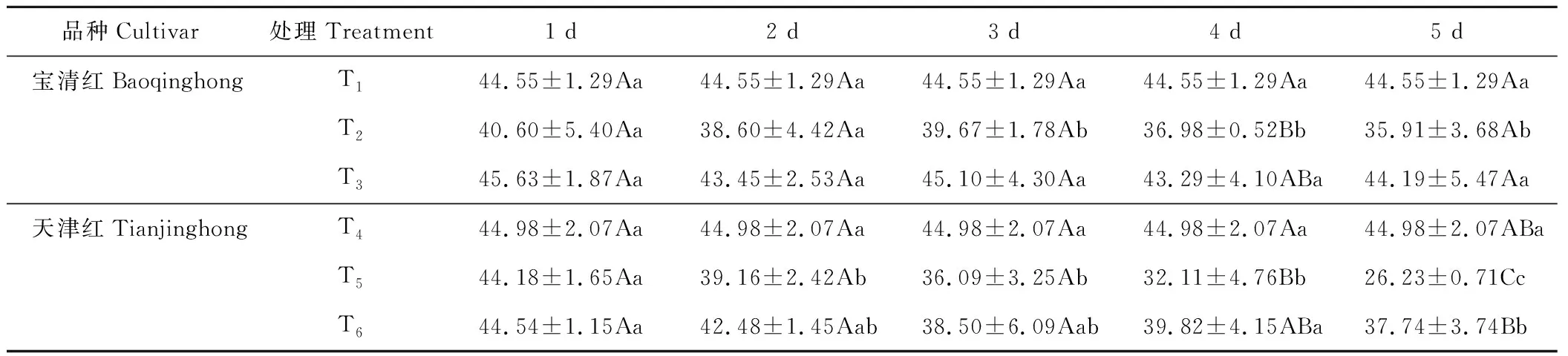
表3 幼苗期低温条件下S3307对红小豆产量的影响Table 3 Effect of S3307 on yield under low temperature stress of adzuki bean during seedling stage (g·pot-1)
3 讨论
S3307是一种重要的植物生长调节剂,在改善植物生长性能、增强植物抗逆性、提高作物产量中发挥着重要作用[20]。S3307通过降低贝壳杉烯氧化酶活性,减少赤霉素(gibberellin,GA)合成前体原料的形成,从而阻抑GA的生物合成,降低内源GA水平,并能够降低生长素(auxin,IAA)水平,同时提高脱落酸(abscisic acid,ABA)水平,前人研究表明不同种类的激素之间存在着相互促进和相互拮抗的生理效应,植物激素间的动态平衡对植物生长发育的调节作用更为重要[21],段娜等[22]指出内源激素在植物生长发育中发挥重要的作用,其含量变化在植物响应非生物胁迫中扮演关键角色。有报道称ABA本身也可以诱导表达ABA合成途径中的酶,从而促使ABA的进一步合成,合成的ABA通过正向反馈机制进一步激活了ABA的大量合成,增强了ABA信号转导途径,从而使植物更好地适应胁迫环境[23]。S3307通过上述的理化功能,使植物体内产生一系列的生理效应,对促进植株抗倒伏、延迟衰老具有明显效果,尤其在作物面对逆境(干旱、低温等)的胁迫时,S3307能显著增强作物对逆境的抵抗能力,从而达到增产的目的[20]。
3.1 幼苗期低温胁迫下S3307与红小豆根系膜脂过氧化和渗透调节的关系
膜系统是受环境胁迫最敏感的部位之一,在植物的抗性生理中具有重要作用。MDA是细胞膜过氧化产物,能加剧细胞膜脂过氧化程度,MDA含量的多少是质膜受损严重与否的重要指标[24],MDA含量与植物耐寒性间呈负相关,胁迫程度越强,MDA积累量越多。游离脯氨酸的积累是植株对逆境的普遍反映,积累的脯氨酸能够降低细胞酸度、稳定生物大分子结构[25],对作物抗逆生长具有较好作用。低温条件下,作物体内积累大量脯氨酸充当能量库来协调细胞氧化还原势的生理生化作用[26]。本试验表明,一定时间范围内,S3307能够明显抑制低温条件下MDA含量的增加,并促进脯氨酸含量增加,这与闫艳红等[27]在大豆苗期的研究结果一致。说明S3307处理能够提高植株的抗氧化能力,保证活性氧清除系统的正常功能,提高了质膜的相对透性,进而提高了幼苗的抗逆性[27]。这可能是因为喷施S3307能够影响植物体内自由基和Δ-二氢吡咯-5-羧酸合成酶(P5C synthetase,P5CS)的合成。自由基作用于脂质发生过氧化反应,最终产物是MDA,喷施S3307抑制了自由基的合成,进而降低了MDA的合成量,增加了作物抗冷性。
3.2 幼苗期低温胁迫下S3307调控红小豆根系抗氧化酶系统
抗氧化酶是指植物体内存在的一系列可以防止自由基对植物造成毒害的具有清除活性氧自由基功能的酶,如SOD、POD、CAT等[28]。逆境条件下,植物细胞产生大量的自由基,如O2·-、OH·、H2O2等,可造成细胞膜脂过氧化反应,进而导致膜系统受到损伤使得作物受到伤害[29]。植物通过SOD、POD和CAT协同作用,使体内的氧自由基维持在较低水平,可在一定程度上减缓或防御低温胁迫[30]。逆境条件下,S3307可有效增强作物的抗氧活性酶活性,进而提高抗逆性和产量[31]。研究指出植物处于逆境时,细胞内自由基产生和清除的平衡系统受到破坏,植物体内存在清除自由基的多种途径,其中最主要的就是抗氧化酶系统,适当浓度的S3307处理可提高植物体内相关抗逆酶活力,增加植株的抗氧化能力。郑春芳等[17]指出S3307能显著提高低温胁迫下SOD和POD活性,进而提高作物的抗逆能力,减少低温对细胞的伤害。刘丽琴等[32]也指出S3307处理可有效促进逆境胁迫下红小豆根系SOD和POD活性。本试验表明,低温处理后,红小豆根系的SOD、POD和CAT活性呈先升高后降低的变化规律,喷施S3307可显著提高SOD、POD和CAT活性,从而降低了膜脂过氧化程度,减少低温对红小豆根系的伤害。其原因与施用外源S3307调控植株内源激素水平有关,喷施S3307正向调控内源ABA水平增加[33],有利于SOD和POD活性的提高,同时可以显著降低内源GA的含量,引起ABA/GA比例大幅升高,促进作物抗冷性能提高,这与李建英等[34]的观点相同。
3.3 幼苗期低温胁迫下S3307影响红小豆根系可溶性物质含量
逆境条件下,植物为了减缓由胁迫造成的生理代谢不平衡,细胞大量积累一些小分子有机化合物,通过渗透调节来降低水势,以维持较高的渗透压,保证细胞的正常生理功能。低温胁迫下可溶性糖和可溶性蛋白在植物体内会大量积累,可溶性糖通过某些糖代谢途径形成保护性物质,提高植物抵抗低温的能力[35]。有报道指出S3307能提高逆境条件下叶片中可溶性糖和可溶性蛋白含量[36],本试验研究结果与此类似,S3307提高了低温条件下红小豆根系可溶性糖和可溶性蛋白含量,但不同品种之间差异很大,耐冷品种的增加幅度高于冷敏品种,这可能与不同品种对外源S3307的应激反应不同有关,耐冷品种除自我保护能力强外,对外源S3307的适应性更好,通过外源S3307加强维持较为正常的碳代谢生理和同化物运输过程,向根系内输送大量糖类物质,并在根系内促进有关酶活性的提高,以分解不溶性的多糖类物质转化为可溶性糖类物质,进而提高作物的抗冷性。总体来说,S3307能通过提高红小豆根系内可溶性糖和可溶性蛋白来增强作物抵抗逆境的能力,并间接促进作物生长。
3.4 幼苗期低温胁迫下S3307提高红小豆产量
孙福东等[37]指出S3307通过显著增加同化物的积累和运输,促进豆荚的建成,提高了产量。王畅等[38]的研究也指出S3307处理能极显著增加芸豆(Phaseolusvulgaris)单株荚数,显著提高芸豆产量。本研究表明,低温处理可降低红小豆产量,尤其是冷敏品种比耐冷品种产量下降幅度更大;低温条件下S3307处理能够提高红小豆的百粒重且显著提高产量。目前,本试验研究了低温条件下S3307处理对红小豆根系抗寒生理和产量的影响,今后将深入挖掘分子水平的机理,进一步拓宽植物生长调节剂对逆境胁迫的缓解效应的研究。
4 结论
幼苗期低温胁迫引起红小豆根系中SOD、POD和CAT活性增强,促进可溶性糖、可溶性蛋白、脯氨酸和MDA含量的提高,导致红小豆产量下降。叶面喷施S3307能通过显著增加可溶性物质和脯氨酸的含量、降低MDA含量及提高保护酶活性等生理过程来抵御低温并降低伤害,缓解产量的下降。耐冷品种宝清红受低温伤害的程度和产量下降均小于冷敏品种天津红,就产量而言,冷敏品种天津红对S3307的响应显著高于耐冷品种宝清红。
猜你喜欢
中老年保健(2022年3期)2022-11-21 09:40:36
农民致富之友(2020年31期)2020-11-18 08:49:34
农村百事通(2019年24期)2019-12-31 06:11:38
科学与财富(2019年15期)2019-10-21 13:35:48
中成药(2018年8期)2018-08-29 01:28:34
发酵科技通讯(2018年2期)2018-07-06 11:38:46
湖南林业科技(2017年1期)2017-02-06 05:29:00
医学研究杂志(2015年9期)2015-07-01 17:28:24
中国当代医药(2015年9期)2015-03-01 02:02:13
农村百事通(2014年14期)2015-02-09 10:36:18
