
荒漠植物囊果草生物学特性及其种群分布格局的生态适应意义
2019-07-27 01:44:44孙海荣车昭碧陈乙实鲁为华王树林李娜娜辛怀璐
草业学报 2019年7期
孙海荣,车昭碧,陈乙实,鲁为华,2*,王树林,李娜娜,辛怀璐
(1.石河子大学动物科技学院,新疆 石河子 832003;2.省部共建绵羊遗传改良与健康养殖国家重点实验室,新疆农垦科学院,新疆 石河子 832003)
囊果草(Leonticeincerta) 为小檗科囊果草属多年生草本植物,是北疆地区低山带温性荒漠中分布的特有物种,为典型的中亚荒漠类短命植物。该植物花期在每年的4月,果期在5-6月,可利用早春积雪融水和春季降水完成整个生活史,其膨大的块状根茎可积累大量的储藏物质,夏季来临后地上部分迅速枯萎。其花期很短,为15 d左右,果实为近球形的囊状结构,称为果囊,种子则藏于果囊之内。膜质果囊侧面和底部具有明显的网状脉络,而顶部则无网状脉络,成熟果囊脱落后顶部极易开裂,内部种子在果囊滚动时完成传播过程。
近年来,由于温性荒漠草地普遍退化,众多学者对温性荒漠草地植被及植物做了大量的研究工作,包括土壤种子库组成及数量特征[1]、各类植物结实特征[2]、种子萌发和种子性状间的关系[3]、各类植物种子的多型性及其对荒漠环境的适应意义[4]、种子性状和家畜消化道传播之间的关系[5]等方面,并取得了诸多研究成果。但对于囊果草的生物学特性、种子传播特征以及野外种群的分布状态方面的研究非常少,仅见有关基本的生物学特征描述[6]和染色体核型分析[7]的零星报道。而北疆地区低山带温性荒漠作为春秋牧场在整个北疆地区草原畜牧业中起着举足轻重的作用,承载着两次利用的“重任”:家畜在春季牧场上恢复体膘和产羔育幼,在秋季牧场上贮存能量越冬并为繁衍下一代进行配种,是连接夏场与冬场的时空纽带,也是草原畜牧业季节间生产畅通的保证[8]。囊果草隶属于荒漠草地中的杂类草植物群落,以伴生种形式出现,具有生长发育快、光合效率高、繁殖率高和结实性极强的特点[9]。早春气温回升,囊果草种子和地下块状根茎便利用冬春雨雪迅速萌发占据有利的生态位,并可在局部地区形成斑块,且囊果草植株适口性好,各种家畜均喜采食[10],对于缓解早春草场牧草短缺的压力和家畜越冬后的体膘恢复具有重要意义。此外,囊果草种群在一定程度上具有增加草地植被类型的多样性、群落结构的异质性[11]及缓冲单一植物类群内同胞之间的激烈竞争的能力,对于维护草地生态系统稳定性具有重要作用。因此,开展囊果草种子传播方式及其种群分布格局的研究对于揭示该物种种群结构和动态变化以及对退化草地的更新与恢复均具有重要意义。
鉴于此,本研究以囊果草为对象,通过对其生物学特性、种子传播特征和野外种群的分布状态进行研究,以期探究如下问题:1)囊果草基本的生物学特性是什么?2)种子传播行为特征如何,环境条件如何影响种子传播过程以及传播行为和果囊结构有什么关系?3)囊果草野外种群的分布格局如何,在不同空间尺度上其分布格局处于什么状态,不同株龄的个体之间存在什么关系?4)囊果草生物学特性、种子传播方式以及种群分布格局对荒漠环境的适应意义。
1 材料与方法
1.1 研究区概况
试验地点位于新疆石河子市南山低山带的温性荒漠(84°58′-86°24′ E,43°26′-45°20′ N),海拔600~900 m,年降水量为150~200 mm,年均温8~10 ℃,极端高温40 ℃以上,土壤为典型的淡栗盖土,黄土母质,土层深厚。该区域植被组成主要以伊犁绢蒿(Seriphidiumtransiliense)为主体,另广泛伴生有藜科(Chenopodiaceae)、十字花科(Brassicaceae)、百合科(Liliaceae)、毛茛科(Ranunculaceae)、禾本科(Poaceae)、小檗科(Berberidaceae)、牻牛儿苗科(Geraniaceae)等多年生、一年生、短命和类短命植物。分布区气候特征为典型的中亚哈萨克斯坦气候型,春季降水丰沛,加之有丰富的积雪融水补给,使得早春短命、类短命植物层片有较好的发育,夏季气候干燥炎热,降水偏少,而秋季降水增加,由此形成了两个植物多样性和生物量的高峰期。在畜牧业生产上,该区域为主要的春秋放牧地,承受着极大的放牧压力,且近年来由于气候环境恶化而普遍退化。
1.2 研究材料
囊果草果囊在2016年5月中旬采集,采集时保证果囊不变形且不开裂。同时在囊果草果期挖取整个植株,保存在密封袋中,用于观测其基本生物学特性。上述样品采集后均带回实验室于4 ℃冰箱内保存备用。
1.3 研究方法
将野外挖取的囊果草植株带回实验室整理,对全株高度、重量进行测定。然后对单株叶片数量、囊果数、囊果重量、每果囊内种子数、种子百粒重、块状根茎高、块状根茎直径以及块状根茎表面瘤状凸起数量进行测定。以上所有数据的测定结果均采用平均值±标准误(mean±SE)的形式表示。
由于囊果草具有果囊这一特殊结构,推测其果囊从母体脱离后,其位移距离与风力和地表粗糙程度密切相关。因此,为探讨囊果草种子扩散行为特征,在实验室内采用可调式小型鼓风机、泡沫板和覆盖纱布泡沫板来模拟野外微地形以及空气流动条件,地表粗糙程度由泡沫板上覆盖纱布的有无及褶皱数量来界定,风力大小由手持式风速测定仪测定。
野外调查时,在囊果草集中分布区内,设置100 m×100 m的样地,在样地内再设置10 m×10 m的样方,将整个样地划分为100个10 m×10 m的格子,依次用卷尺测定每个格子内囊果草植株的坐标点,然后推算出整个样地内每株囊果草的坐标位置,并根据囊果草单株结果与否,将其分为结果和不结果植株两个类群,结果植株为成株,不结果植株为幼苗,用于种群的空间格局和个体之间的关联特征分析[12]。
1.4 数据处理与统计方法
采用点格局分析方法分析囊果草空间格局[13-14],其计算过程利用ADE4软件包中的Ripley’s K函数以及Monte Carlo随机模拟方法,计算时用Monte Carlo方法模拟999次,空间尺度为0~50 m,步长为0.5 m,采用ADE4软件包中Text To Bin. exe和Bin To Text. exe模块进行二进制和十进制数据的相互转换,进行密度函数和关联度函数计算,获得100个密度函数和关联度函数值,之后利用Sigmaplot 10.0进行各函数曲线图的绘制,依据密度函数和关联度曲线在95%包迹曲线的位置判定种群的分布状态和不同囊果草类群的关联程度。
采用Excel初步整理基本生物学特性数据,并利用SPSS 19.0进行数据的单因素方差分析(One-way ANOVA)以检验结果和不结果植株各性状的差异性,采用Duncan多重比较法。采用回归分析来阐明果囊传播距离大小和果囊体积之间以及不同风力和地表粗糙程度与果囊传播距离大小之间的关系。
2 结果与分析
2.1 囊果草基本生物学特性
囊果草为多年生草本,有果植株高(188.09±33.03) mm,无果植株高(162.02±30.60) mm。结果单株重量(16.24±6.48) g,不结果单株重量(6.21±0.50) g。基生叶2~3枚,2~3回三出深裂,结果植株叶的裂片数(13.91±2.54)枚,不结果植株叶的裂片数量(5.23±3.28)枚。
总状花序顶生,长4~6 cm,具总梗,长2.0~2.4 cm,无毛;萼片6,花瓣状,椭圆形或卵形,黄色,外侧带有蓝紫色斑,长于花瓣;花具有暂时闭合特性;花瓣蜜腺状,倒卵形,基部呈爪;雄蕊6,分离,花药顶部2瓣开裂;心皮1,胚珠2~4枚。
结果植株每株囊果数(8.29±4.82)个,每个果囊内种子数(2.82±0.61)个,囊果重量(0.20±0.10)g,果囊膨胀成膀胱状,侧面和底部具明显的棕色网状脉,而顶部薄且无网状脉但具蓝紫斑,成熟时易破裂,种子球形,藏于果囊内。
块状根茎近似卵状、球状或不规则,结果植株块状根茎高(34.50±10.19) mm,直径(23.09±6.51) mm,表面密布瘤状凸起,平均数量为(6.11±2.84)个;不结果植株块状根茎高(23.35±6.43) mm,直径(18.09±6.43) mm,表面瘤状凸起较少,平均数量为(0.60±0.14)个,且不同株龄块状根茎差异极大,所有株龄植株须根少且入土浅。两类植株基本生物学特性见表1和图1。
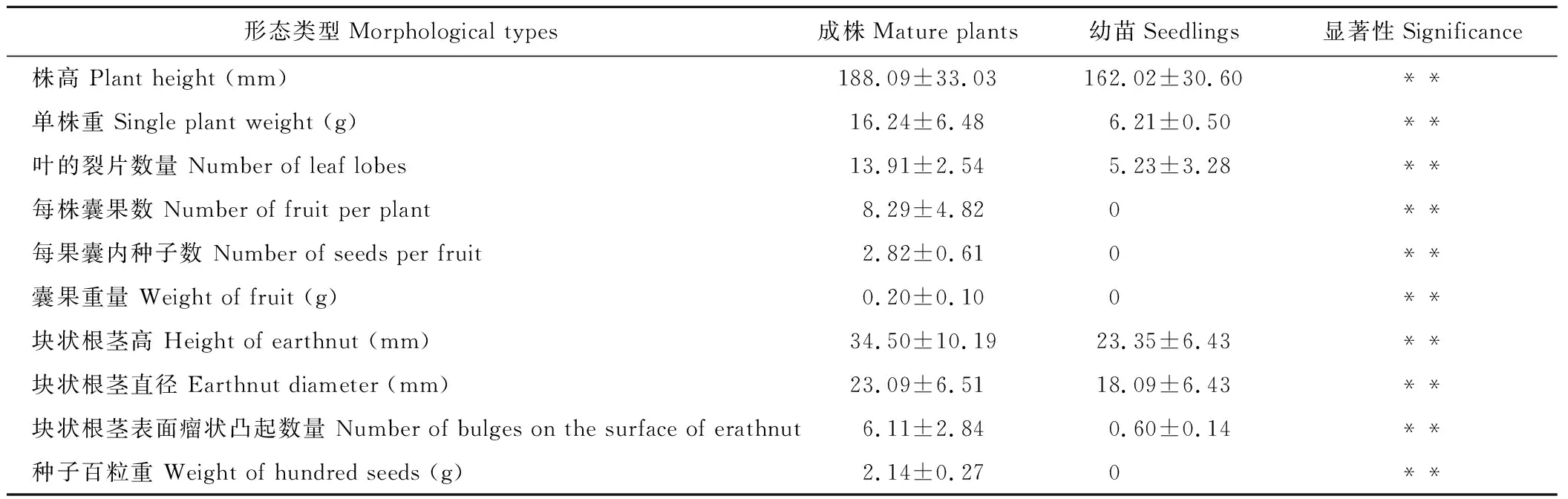
表1 囊果草基本生物学特性Table 1 Biological characteristics of L. incerta
注:**表示同一指标不同处理间在0.01水平差异显著。
Note:** indicate the same index between different processing significant at 0.01 level.

图1 囊果草植株(A)、块状根(B)及果囊(C)形态特征Fig.1 Morphological characteristics of L. incerta seedling (A), tuberous roots (B) and fruits (C) A: 1. 幼苗Seedling; 2. 花期植株Flowering seedling; 3. 果期植株 Bearing fruit seedling; 4. 成熟后顶部开裂的果囊 Dehiscent top of fruits;B: 不同株龄的块状根茎形态Morphological characteristics of tuberous roots;C: 1. 果囊底部 Bottom of fruit; 2. 果囊侧面 Profile of fruit; 3. 果囊顶部 Top of fruit.
2.2 囊果草果囊传播及果囊内种子散布特征
果囊体积大小是影响囊果移动距离的重要因素,通过对不同微地形条件下囊果移动距离与果囊大小的关系进行模拟实验,并对实验结果进行回归分析。结果表明,在不同粗糙程度的微地形下,随着果囊体积的增大,囊果移动距离呈线性递增的趋势,且相关系数均达到显著水平(P<0.05)(图2),表明拟合结果可信。说明在不同地表粗糙度的微地形条件下,较大体积的果囊有助于囊果的远距离移动。

图2 果囊大小(体积)与囊果传播距离的关系Fig.2 The relationship between fruit volume and fruit dispersal distance A: 光滑表面 Smooth surface; B: 平铺纱布Tiled gauze; C: 稀疏褶皱 Sparse drape gauze; D: 密集褶皱 Intensive drape gauze.
地形特征与果囊内种子的散布特点密切相关,通过控制平板表面纱布的有无以及纱布褶皱的疏密来模拟地表粗糙程度对果囊内种子散布的影响,结果表明,随着地表粗糙程度的增加,种子最短散布距离显著降低(P<0.05),而每个果囊内散布的种子数量显著增加(P<0.05),当平板表面光滑时,种子散布最短平均距离为151.76 cm,当平板表面铺有密集褶皱纱布时,种子散布最短平均距离显著降低至78.50 cm (P<0.05);而当平板表面光滑时,每个果囊散布的平均种子数量为1.16个,当平板表面铺有密集褶皱纱布时,每个果囊散布的平均种子数量显著增加至1.97个(P<0.05)(图3)。果囊移动距离与种子散布数量之间具有显著负相关关系(r=0.30,P<0.05),随果囊移动距离的增加种子散布数量呈现增高的趋势。
2.3 囊果草种群分布格局及种内个体间关联分析
点格局分析结果表明,囊果草种群包括幼苗和成株在0~2 m尺度上均表现为均匀分布,2.5~14.0 m尺度上表现为聚集分布,在大于14 m的尺度下表现为随机分布(图4A);囊果草成株在0~2 m尺度上表现为均匀分布,在2~11 m尺度上表现为聚集分布,在大于12 m尺度上表现为随机分布(图4B);囊果草幼苗在0~2 m尺度上表现为均匀分布,在3~12 m尺度上表现为聚集分布,在大于12 m尺度上表现为随机分布(图4C)。综合以上分析结果可知,随着空间尺度的扩大,囊果草种群的分布特征表现为从均匀分布到聚集分布再到随机分布的特点。
通过对囊果草成株和幼苗的空间关系的关联分析,结果表明在0~2.5 m尺度范围内二者表现出负关联,但关联性不显著;在3.0~17.5 m尺度下表现为正关联,且关联性呈现先增高后降低的过程,在8 m时达到最大;在大于18 m的尺度范围内,囊果草的成株和幼苗逐渐由负关联性向正关联性过渡,但关联性均不显著(图4D)。从囊果草幼苗和成株的空间关联来看,正关联仅存在于中等尺度上3.0~17.5 m,随着空间尺度的递增,二者之间的关联性逐渐减小且关联性不显著。

图3 地形特点与种子散布距离和数量的关系Fig.3 The relationship of terrain condition and seed dispersal distance and the number of dispersed seeds 不同小写字母表示不同处理间种子散布最短距离与每果囊散布种子数差异显著(P<0.05)。 Different lowercase letters indicate the shortest between different treatments and the number of seeds per fruit sac significant difference at 0.05 level.

图4 囊果草种群空间分布格局及种内幼苗和成株关联特征Fig.4 Spatial distribution patterns of L. incerta population and spatial association characteristics of seedlings and mature plants A: 所有植株空间分布格局Spatial patterns of L. incerta population; B: 成株空间分布格局Spatial patterns of mature L. incerta population; C: 幼苗空间分布格局Spatial patterns of immature L. incerta population; D: 成株和幼苗的个体关联Spatial association of two L. incerta population.L(r): 密度函数值 Density function value; M(r): 关联系数 Correlation coefficient.
3 讨论
3.1 囊果草形态特征与生态适应性的关系
囊果草属于高山荒漠特有植物,且其物候期较短,较难发现,所以对于囊果草的生物学特性、种子传播特征以及野外种群的分布状态方面的研究仍然处于基础阶段,仅见有关基本的生物学特征描述[6]和染色体核型分析[7]的零星报道。干旱少雨和蒸降比悬殊是干旱荒漠的显著特点,维持水分平衡是保证旱生植物生存繁育的首要条件[15],干旱荒漠区的植物能凭借特殊的适应机制顽强的生存和繁衍。囊果草具膨大的块状根茎,是植株营养物质和水分的主要贮存部位。成年植株块状根茎直径可达23 mm,块状根茎表面生有须状根系和瘤状突起,这一结构有助于在降水期迅速、高效地捕获土壤浅层的水分并及时地储存在块状根茎中,以备旱期利用[16]。囊果草植株地上部分整体呈现圆锥形,有利于光合作用。且囊果草株高较低矮,在160~180 mm左右,与块状根相比在整株重量中所占比例较小,这种分配策略有助于提高植株的根茎比[17-18],对植株在干旱环境中通过增加同化物向根的转移和扩大水分的吸收量具有重要意义[19]。囊果草的茎秆和叶片均为肉质型,在贮存养料和水分的同时,还具有一定的韧性,且叶片几乎贴于地面,茎秆高度在种子成熟后恰好被果囊完全利用,可保证植株耐受强风地吹蚀,不易折断[20];且叶片边缘卷合,这一特性能使有效叶面积减小,减缓因叶面蒸腾而导致的水分亏缺,有利于适应干旱缺水的环境[21],叶片表面附有的白色蜡质层在缓解叶片蒸腾失水和避免太阳辐射等方面发挥重要作用[22]。在荒漠生态系统中,水分胁迫是限制荒漠植物生长的主要因子[23],囊果草为了适应特定的荒漠生境条件,在长期的进化过程中形成了一套完备的耐旱机制,包括具有膨大的块状根茎、肉质的茎叶、较大的根茎比和特殊的叶片结构等,其主要目的是减少蒸腾、保持水分[24-25]。
荒漠生态系统的另一大特点是昼夜温差大,通过野外观察发现囊果草的花具有温度变化导致的花暂时性闭合特性,这一特性对于囊果草在多变的荒漠生境中进行繁育具有重要意义。因为花暂时性闭合的特性在促进自主自交的同时,还能保持花内的微环境温度,提供较稳定的花微环境温度可促进花粉萌发率及柱头可授性;在下雨前通过花暂时性闭合来避免雨水对花粉的冲刷以及对花粉活力和柱头可授性的影响;在大风前的花暂时闭合性通过保护花粉聚集水平来影响传粉者的传粉效率或促进自主自交率[26]。囊果草花的暂时性闭合是该物种在长期适应早春环境中,为了促进自主自交、延长雌雄器官的功能及提高繁殖成功而进化出的特殊适应机制。
3.2 囊果草的种子传播特性
种子传播是指种子以某种方式离开母株到达适宜生境萌发成苗实现定居的过程[27],种子传播也是植物迁移到不同生境的主要手段,种子迁移到远离母株的地方,减少了种群内部的竞争,有利于整个种群的生存繁衍及拓展生存空间,这是植物在漫长的演化过程中形成的对环境的适应能力[28]。本研究发现在不同粗糙程度地面上囊果草果囊的滚动距离随果囊体积大小的增加均呈线性递增趋势,表明较大的果囊有助于携带囊果内部的种子实现远距离传播。对荒漠区藜科植物的种子传播特性进行研究表明,散布体扩大面积不是为了在空中飞翔或滑翔,而是为了在地表滚动,滚动过程中释放出种子,这种植物称为风滚植物[29]。囊果草是典型的风滚式传播植物,较大的球形果囊是一种附属结构,不过该结构并不像具种(果)翅传播体那样可以借助发达的种翅使种子“御风飞行”[30],而是为了在地表风滚传播时增大果囊的受力面积,使果囊借助风力滚向远方。值得注意的是,囊果草成熟果囊脱落后,随风滚动过程中将内部的种子散布出来,球形种子流动性较好,脱离果囊后仍可借助微地形变化进行小尺度移动,直到遇见地表裂缝或坑洼处,种子沉降实现定居,即囊果草种子有二次传播特性[31]。对风滚植物角果藜(Ceratocarpusarenarius)种子传播特性的研究发现角果藜植株在随风滚动时,植株残体上的种子随植株的滚动四处传播,从而扩大了种子的传播范围[32],这一传播特性和囊果草类似。进一步研究表明,囊果草的果囊在滚动过程中,随着地表粗糙程度的增加,种子散布的最短距离显著减小,而果囊内部释放的种子数量则显著增大,说明粗糙坎坷的地形可使囊果草种子提前从果囊内释放,另一方面,囊果草膜质果囊结构特殊,果囊底部和侧面均具有网状结构脉络,且质地坚硬,不易破损,而顶部的囊膜没有硬质脉络结构,在粗糙不平的地面上滚动时由于颠簸极易破损,从而释放果囊内部的种子,且地表越粗糙,释放的种子数量越多。有研究表明角果藜种子在风滚传播时,随着植株移动距离的增加,种子传播数量逐渐减少[32],这与本研究所得结果一致。
值得注意的是,囊果草为多年生植物,地上植株部分在夏季完成种子传播过程以后迅速枯萎,膨大的块状根茎在地下以休眠方式度过不良环境,以待来年环境条件适合时发芽长成新植株。一方面,地上种子随囊果进行风滚式传播,使植物的后代逃离不利的环境,寻找比较适宜的环境[33],属于“高风险”的“逃避策略”,有利于种群的迅速增长与扩张[34];另一方面,地下块状根茎使植株在原位萌发[35],为种群在原分布区的繁衍提供了可能,使其能够在恶劣的荒漠环境达到最大繁殖成功率[36],属于“保护型”的“谨慎策略”,分散了植物生存的冒险性[37]。青藏高原的特有植物块茎堇菜(Violatuberifera)也采用类似的繁殖策略[38]。囊果草的生殖策略,增加了子代的存活机会,减小了环境的时空变化对植株生殖成功的影响,允许植物在高度不稳定的环境中成功定居[39],这是囊果草适应荒漠多变环境的一种生存方式。
3.3 囊果草种群的分布格局
种子散布是植物生活史中的重要阶段,影响着种子时空尺度上的定居方式、种子与母株的距离、种子到达的生境类型以及建成的植株将与何种植物竞争,从而影响种子和幼苗的存活,最终影响母株及后代植物的适合度和种群结构[40]。通过点格局分析方法对比囊果草种群分布格局及种内个体间关联性进行研究,结果表明,随着空间尺度的增加,囊果草种群包括成株和幼苗的分布特征均表现为从均匀分布到聚集分布再到随机分布的特点,这种分布特征与其繁殖方式密切相关。在小尺度范围内(0~2 m),囊果草依靠地下块状根茎的发芽形成成株,形体较大的植株由于种间竞争效应使得种群有倾向于均匀化的趋势[41-42]。囊果草形体较小的幼苗由种子萌发所得,而种子传播数量随果囊移动距离的增加呈现增高的趋势,同时形体较小的植株发生种内竞争的可能性很小,密度制约效应在短时间内还未发生,因此在中等尺度范围内(3~12 m)幼苗表现聚集分布。通过对伊犁绢蒿种群空间分布的特点的研究也得出类似结论[2]。
从囊果草成株和幼苗两个群体的空间关联来看,正关联仅存在于中等尺度上(3.0~17.5 m),随着空间尺度的递增,二者之间的关联性逐渐减小且关联性不显著,这说明不同种群的关联性只发生在中等尺度范围内,由此可见种群内部关系对空间尺度具有强烈的依赖性[43],形成这种分布格局的主要原因和囊果草种子扩散、萌发以及幼苗建成有关,因为在中等尺度范围内囊果草每个果囊所传播的种子数量最多,若是在此尺度范围内种子恰好落在成株旁边,成株较大的地上冠层会给幼苗以适度的庇护作用,在成株庇护之下的个体存活的概率较大[44],因此这种关联性只发生在中等尺度范围内。有研究表明,伊犁绢蒿的幼苗和成株这两个种群的空间关联性也遵循这一规律[43]。由此可见,囊果草通过地上种子风滚传播和地下块状根茎营养生殖对自身种群结构以及种群内不同个体间的空间关联性产生影响。
猜你喜欢
内蒙古民族大学学报(社会科学版)(2020年2期)2020-11-06 09:08:52
今日农业(2020年13期)2020-08-24 07:35:28
中成药(2017年3期)2017-05-17 06:09:00
中国神经再生研究(英文版)(2017年4期)2017-01-12 02:48:06
太空探索(2016年5期)2016-07-12 15:17:55
当代化工研究(2016年7期)2016-03-20 16:21:59
文苑(2015年5期)2015-05-11 10:47:30
电子设计工程(2015年8期)2015-02-27 12:05:34
时代英语·高三(2014年5期)2014-08-26 17:01:17
天然产物研究与开发(2014年3期)2014-04-27 14:15:29
