
黄野螟成虫触角电镜观察及其对7种杀虫剂的EAG反应
2019-07-25 02:28赵鹏飞常明山吴耀军方小玉杨振德周红英
广西林业科学 2019年2期
赵鹏飞,常明山,吴耀军,罗 辑,方小玉,杨振德,周红英
(1.广西壮族自治区林业科学研究院 广西优良用材林资源培育重点实验室,南宁 530002;2.广西大学林学院,南宁 530004;3.崇左市森林病虫害防治检疫站,广西崇左 532200;4.广西国有高峰林场,南宁 530001)
黄野螟(Heortia vitessoides)属于鳞翅目螟蛾科(Pyralidae),是土沉香(Aquilaria sinensis)生长过程中最重要的食叶性害虫[1]。在国内主要分布于广东、广西、云南、福建、海南和台湾等地,国外主要分布于印度、斐济和尼泊尔等国家[2]。初孵幼虫聚集危害叶片,被咬食后的叶片仅剩一层网状叶脉和叶表皮,低龄幼虫在叶片之间的转移有跟随的特点。四龄幼虫开始逐渐分散在不同叶片,暴发时可以将整片叶吃光,导致整株树枝被吐丝包裹。高龄幼虫啃食后的树干嫩梢和枝条极易干枯,林木光合作用受阻,难以生长,持续危害至整株死亡。该虫在我国南方地区每年发生多代,成虫产卵量大,危害十分严重,是土沉香种植过程中需要重点监测和防治的害虫。
我国对螟蛾科昆虫触角感受器的研究主要集中在玉米、竹类等[3]的相关昆虫。张蒙等[4]研究报道了黄野螟幼虫触角和口器上的感器分布和类型。由于黄野螟成虫多在土沉香叶梢、叶背产卵,说明成虫的感受器官能够对产卵的位置进行有效定位[5]。成虫触角是感受寄主信息化合物的重要器官,因此成虫触角上感受器的分布、类型和数量的差异将直接影响该过程。杀虫剂利用化学物质影响昆虫的感受器官和搜索行为,李会仙[6]报道高效氯氰菊酯对鳞翅目棉铃虫有明显驱避和拒食作用。杀虫剂还会干扰昆虫定位行为,影响昆虫交配和产卵等行为[7]。本试验通过观察成虫触角的超微形态、结构和分布,描述两性的触角种类、形状和差异,选取7种杀虫剂测定黄野螟成虫触角电生理反应,为开展黄野螟成虫触角化学感器的感受功能研究以及探索杀虫剂对黄野螟成虫行为干扰提供科学依据。
1 材料与方法
1.1 试验材料
本试验所用黄野螟幼虫均采自广西凭祥市东盟林业产业园种植区的土沉香林。将采集的幼虫放入网袋内,采集500头以上,带回室内于人工气候箱内饲养,喂以新鲜土沉香叶片,待养至成虫后,喂食15%的蜂蜜水补充营养。饲养温度为(26±1)℃,相对湿度为85%,光周期为16∶8(L∶D)。每次试验分别取新羽化的雌雄成虫进行扫描电镜观察和触角电位检测。
1.2 试验药剂
根据不同杀虫剂对成虫、蛹和高龄幼虫的试验结果及毒杀机理选择苏云金杆菌(Bacillus thuringiensis)500倍液(美国杜邦公司)、乙基多杀菌素1 000倍液(美国陶氏益农公司)、34%乙多·甲氧酰肼1 500倍液(美国陶氏益农公司)、20%氯虫苯甲酰胺3 000倍液(美国杜邦公司)、球孢白僵菌2 000倍液(北美联邦农大集团)、灭幼脲200倍液(美国陶氏益农公司)、绿僵菌2 000倍液(北美联邦农大集团)和3%苯氧威200倍液(北京鑫敦丰公司)。选择去离子水作为对照处理(CK)。
1.3 试验方法
1.3.1 触角电镜扫描法
触角感器检测方法参照孟倩倩等[8],略作改进。采用S-3400N扫描电子显微镜(日本日立公司),加速电压为5.0 kV。取新羽化的黄野螟雌、雄成虫各5头,放入-20℃冰箱中冷冻10~15 min,待成虫昏迷后,将带有完整触角的成虫头部切下,依次用梯度乙醇脱水20 min,将雌、雄黄野螟成虫触角依次切下,在室温下风干,涂一层双面胶于干燥的电镜样品台,将触角均匀地粘贴在样品台上,喷金膜(20 nm)后放入电镜。观察触角的感器类型、长度和分布等,并拍照。
1.3.2 触角电位测定法
触角电位(Electroantennography,EAG)是昆虫化学生态学中重要的电生理技术,采用触角电位仪(德国SYNTECH公司)进行记录和测定。取新羽化的雌、雄黄野螟成虫各5头,用锋利的刀片将黄野螟触角从基部切下,鞭节末端切除2 mm,用导电胶将其横搭在电极两端,随机抽取不同杀虫剂进行测试。在测定过程中,触角的活性会因试验时间的增加而降低,在试验中标定触角的活性,在每一杀虫剂(包括各种浓度)EAG活性测定的前后均做空白标样的EAG测定[9]。触角电位分析所用数据均为校正后的EAG反应相对平均值,即将样品EAG测量值减去标样处理值后得到的平均值。刺激气流流速、持续气流均设定为400 mL/min,刺激时间0.5 s,2次刺激间隔60 s。将剪成2 cm×0.5 cm的定性滤纸纸条折成V字形作为每种溶液的载体,测试剂量为10 μL,将触角和药剂分开放置。各种测定每根触角重复3次。以去离子水作为对照(CK)。
2 结果与分析
2.1 触角的结构特征
黄野螟成虫触角的外观形态上略有差异,雄虫触角的鞭节亚节略多于雌虫。触角均为线状,由1节柄节、1节梗节和70~75个亚节组成(图1),触角长9 300~9 800 μm,柄节粗大,梗节较短,延长呈毛状;柄节长约300 μm,最宽处约400 μm;梗节长约100 μm,最宽处约260 μm;柄节、梗节两侧密被不规则的鳞片,鳞片细长,呈扇形;鞭节长8 800~9 100 μm,是表面感器类型最多的节。触角背面有层次分明的鳞片,层层相叠,少有感器分布。

图1 黄野螟触角和鞭节微观形态Fig.1 Microscopic morphology of antennae and flagellum of Heortia vitessoides
2.2 感受器的类型和特征
黄野螟成虫触角表面具有6种触角感器:毛形感器,鳞形感器,腔锥形感器,刺形感器,耳形感器,栓锥形感器(图2)。
2.2.1 毛形感器(Sensilla trichodea,Str)
毛形感器是黄野螟成虫触角上数量最多、分布最广的感受器。触角上的毛形感器有2种:毛形感器Ⅰ型长49~58 μm,直径1.5~3 μm,基部较粗,由基部向顶端逐渐变细,顶端弯曲;基部斜生,与表面形成钝角,着生于表皮皱褶形成的蜂窝状臼状窝内,规则排列;全感器呈弧形生长,感器壁上有螺纹。毛形感器Ⅱ型长23~29 μm,直径1.5~3 μm,顶部不弯曲,不同于毛形感器Ⅰ型。两性间差异较大,黄野螟雄虫触角鞭节的毛形感器在数量上多于雌虫,且雄虫的毛形感器较长。
2.2.2 鳞形感器(Sensilla squamiformia,Ssq)
鳞型感器长60~70 μm,臼状窝直径2~3 μm。分布于鞭节鳞片周围,比鳞片窄,形状如触角表面覆盖的鳞片,中间宽,两端细。基部着生在臼状窝内,数目较少,仅分布在鞭节的2个亚节的连接处,雌雄感器无明显差别。
2.2.3 腔锥形感器(Sensilla coeloconica,Sco)
腔锥形感器分布于黄野螟成虫触角鞭节的腹面,分为2种类型:ScoⅠ 长9.5~10 μm,直径5.6~6 μm,由外部圆形腔和内部圆形腔中心直立的柱状栓2部分组成,圆腔四周没有缘毛。ScoⅡ长 5.5~6 μm,直径 4.5~5 μm。形状像一朵花蕾,是触角表皮向下凹陷而形成的一个圆形腔,圆腔中央着生一明显中心锥,中心锥呈圆锥形,整体内卷,周围分布着16~18个缘栓,这些缘栓逐渐向中间聚拢生长,雌雄感器无明显差别。
2.2.4 刺形感器(Sensillum chaetica,Sch)
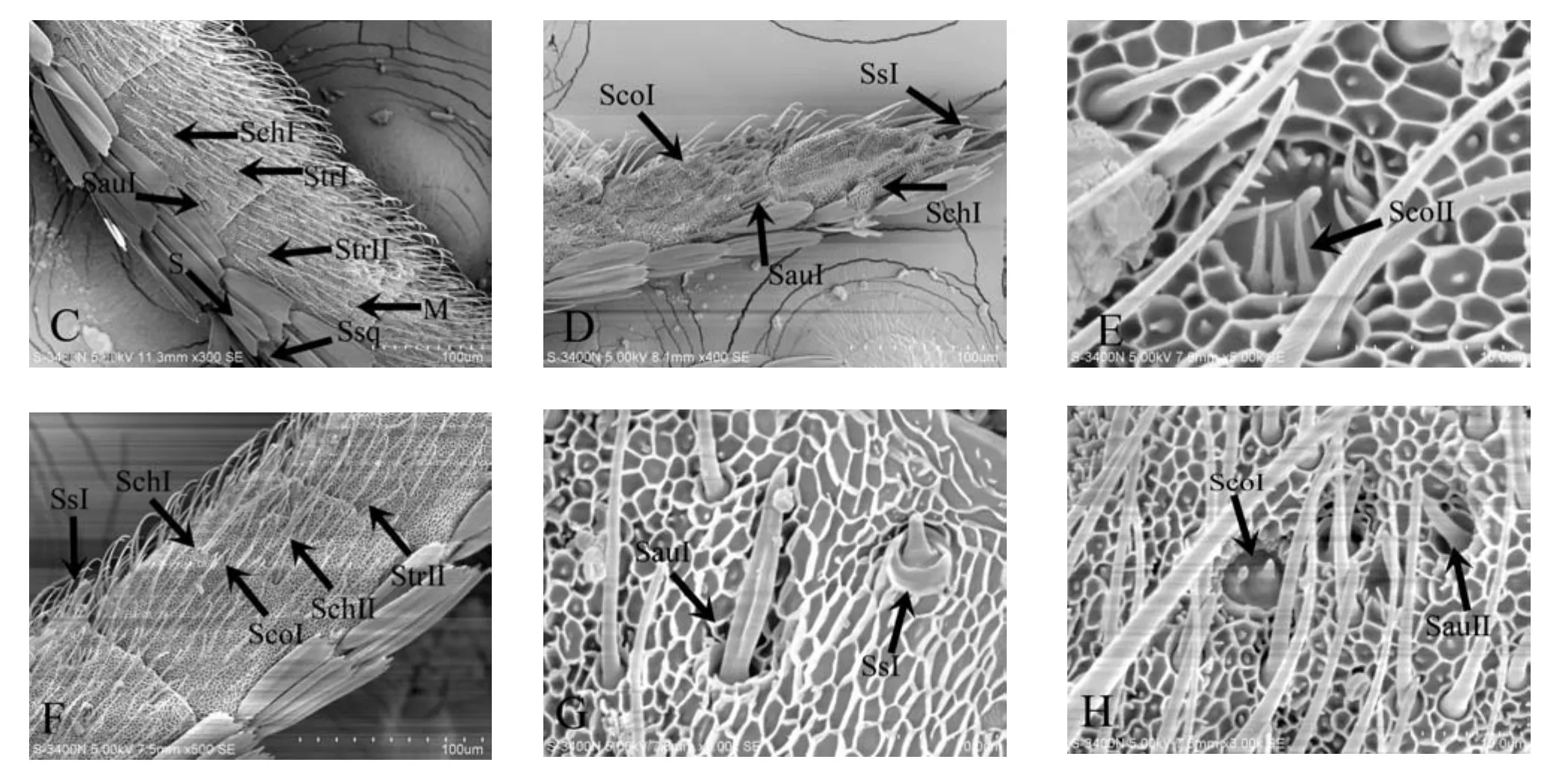
图2 黄野螟成虫触角上的各种感器Fig.2 Sensilla types in antennae of Heortia vitessoides adults
雌虫的刺形感器长42~55 μm,直径2.0~2.5 μm;雄虫为55~65 μm,直径4 μm。I型感器着生在触角表皮特化形成的臼状窝内,笔直斜生,与毛形感器区分明显。臼状窝生长在触角表面的蜂窝状网纹窝里,多分布于鞭节中央和末端位置;刺型感器Ⅱ型长50~55 μm,直径4 μm,跟Ⅰ相似,在基部以上发生分叉,Ⅱ型未在雌虫触角发现。
2.2.5 耳形感器(Sensilla auricillica,Sau)
耳形感器较毛形感器粗短,分布于黄野螟成虫触角上,主要有I型和II型2种,着生于鞭节交接处。I型耳形感器长约22~25 μm,直径1.5~2.5 μm,贴靠触角腹部生长,形似心叶状,顶端钝圆,基部以上部分中部凹于两侧;II型耳形感器长约8~9 μm,直径1.5~2 μm,舌形,着生于表皮内凹形成的圆形腔内,整体垂直于表皮生长,雌雄感器无明显差别。
2.2.6 栓锥形感器(Sensilla styloconica,Ss)
除端节外,每节前缘均着生有栓锥形感器。长度28~30 μm,直径8.5~9 μm,是最大的感器之一。整个感器呈拇指状,感器上具有网纹,仅端部锥状处无覆盖。鞭节末端着生数量为3个,呈品状分布。
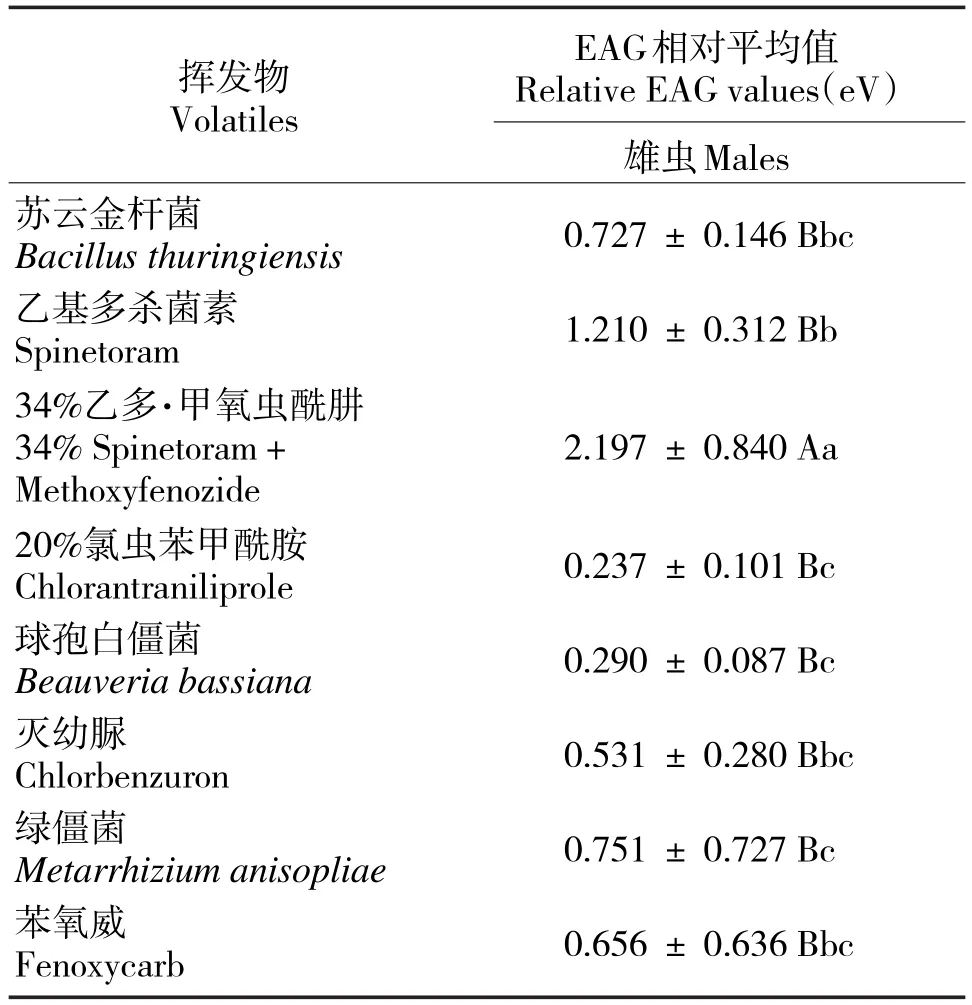
表1 黄野螟雄成虫触角EAG相对平均值Tab.1 Relative mean value of tentacles EAG of male Heortia vitessoides adults
2.3 黄野螟成虫触角对杀虫剂的反应试验
黄野螟雄虫对乙多·甲氧虫酰肼的反应相对平均值最大(2.197 eV,P<0.01),呈极显著性差异;对乙基多杀菌素的反应相对平均值较大(1.210 eV,P<0.05),呈显著性差异;对球孢白僵菌、氯虫苯甲酰胺和绿僵菌的反应相对平均值较小(表1)。
黄野螟雌虫对苏云金杆菌的反应相对平均值最大(1.601 eV,P<0.01),呈极显著性差异;其次为乙多·甲氧虫酰肼(0.895 eV,P<0.05),呈显著性差异;对绿僵菌和氯虫苯甲酰胺的反应相对平均值较小(表2)。
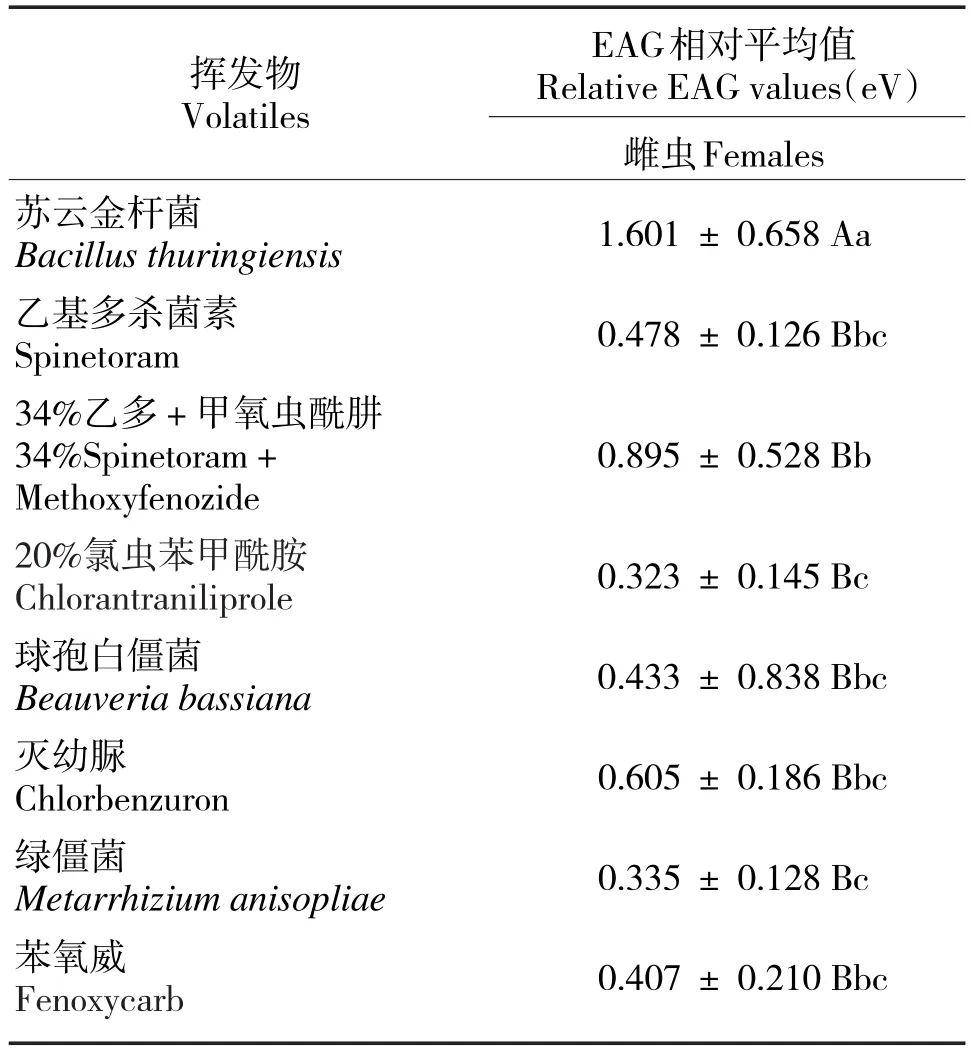
表2 黄野螟雌成虫触角EAG相对平均值Tab.2 Relative mean value of tentacles EAG of female Heortia vitessoides adults
3 讨论与结论
本研究对黄野螟雌雄成虫触角进行扫描电镜观察,发现黄野螟触角分布6种感器:毛形感器、鳞形感器、刺形感器、腔锥形感器、耳形感器和栓锥形感器。其中毛形感器、刺形感器、腔锥形感器和耳形感器均有2种类型。大部分感受器着生在鞭节腹面,这一特征与已报道鳞翅目昆虫一致[12-15]。其中鳞形感器、腔锥形感器和耳形感器属于在黄野螟触角微观结构中首次发现,杨毅等[10]报道的黄野螟的柱形感器和钟型感器未在此研究中发现。
黄野螟雌雄成虫触角的形状基本相同,但两者在感器的种类、形态和数量上存在一定的差异。有关黄野螟的触角感器的性二型性,本研究发现黄野螟雄虫触角鞭节的毛形感器在数量上多于雌虫,雄虫的毛形感器长于雌虫。这和刘志雄等[11]的报道雄蛾毛形感器数量大于雌蛾,雄性昆虫对于性信息素更加敏感一致;在黄野螟触角中发现Ⅱ型刺型感器只在雄虫触角上分布,雌虫触角上未见此感器类型;此外还发现雄虫的栓锥形感器数量明显多于雌虫。虽然昆虫的触角种类和功能不完全相同,但是同一类触角感器基本形态和触角感器种类有一定的相似性。黄野螟触角鞭节表层呈网状纹结构,这与已报道过的豆野螟和草地螟等螟蛾触角表面结构相似[12-13]。毛形感器呈毛发状,分布较广,可能是其感受信息化合物的主要感器[14]。黄野螟的鞭节顶端均无毛形感器,着生刺型感器,刺形感器钢直如刺,可能具有感受机械刺激的功能[15],昆虫的刺形感器对于求偶微环境以及寻找适宜的觅食和居住场所等具有选择行为功能[16-17]。栓锥形感器的主要作用可能是感受寄主植物挥发物和外界气味因子,能够感受信息素等[18]。耳型感器在其行为倾向和生理学特性方面发挥重要作用,对声波有反应,可能为物理感受器[19];腔锥形感器也具有感受植物挥发物的功能[20]。
黄野螟由于繁殖迅速,危害严重,存在世代重叠的现象,杀虫剂会对其整个生活史产生影响。黄野螟成虫接触到杀虫剂后有微妙行为变化,可能是其应对杀虫剂的预警信号。在触角电位检测中,雌成虫对苏云金杆菌的反应相对平均值最大;对乙多·甲氧虫酰肼的反应相对平均值次之。在化学防治的过程中,杀虫剂会对周边环境造成影响,干扰触角感受外部化学信息进行准确的定位,赖敏[21]报道了氯氰菊酯和溴氰菊酯对瓜实蝇雌虫具有较好驱避作用和产卵抑制作用,溴氰菊酯对瓜实蝇雌虫的驱避作用最强,驱避率为83.33%;黄野螟雄成虫对乙多·甲氧虫酰肼的电生理反应相对平均值最大,有极显著性差异;对乙基多杀菌素的电生理反应相对平均值较大,有显著性差异。黄野螟成虫在求偶和选择正确的寄主植物器官产卵过程中,也会通过触角接收来自外界的化学信号。宋亮[22]报道小菜蛾成虫在施药叶片上的产卵率显著下降,这可能与杀虫剂对小菜蛾成虫的刺激作用有关。通过研究杀虫剂对黄野螟成虫触角的刺激反应,可为深入研究杀虫剂对黄野螟成虫行为干扰提供理论依据。
本研究仅对黄野螟触角的微观结构及其对7种杀虫剂触角电生理反应进行研究分析。植物挥发物和昆虫信息素等也能对昆虫的取食、产卵和趋避等产生影响,有关上述化合物之间的互作反应有待进一步研究。
猜你喜欢
热带生物学报(2022年6期)2022-11-29
生物学杂志(2021年4期)2021-08-24
昆明医科大学学报(2021年1期)2021-02-07
环境昆虫学报(2020年5期)2020-11-11
河南农业科学(2019年5期)2019-05-28
中国水稻科学(2017年2期)2017-04-05
河北林业科技(2016年5期)2016-11-08
中华老年口腔医学杂志(2016年5期)2016-03-01
中国继续医学教育(2015年5期)2016-01-07
天津大学学报(自然科学与工程技术版)(2015年10期)2015-12-29
