
花生种子发芽期呼吸酶活性对淹水的响应
2019-06-18 11:27瑊刘登望李林蒋薇刘振宏
花生学报 2019年4期
湛 瑊刘登望李 林蒋 薇刘振宏
水分是影响农作物生长发育、区域分布和生产水平的最重要生理生态因子之一,而渍涝是世界作物生产的主要非生物逆境胁迫之一。在降水量较大、水位波动剧烈或排水设施较差的地区常会遭遇渍涝的危害。据统计,全球10%的灌溉耕地受到渍涝灾害影响,导致农作物产量损失高达20%[1-2]。近50年来,我国农作物多年平均洪涝灾害受灾面积约占全国播种总面积的6.4%,约为920万hm2,其中长江中下游地区和黄淮海平原受渍涝灾害的影响面积最大,约占全国总受灾面积75%以上[3-4]。随着全球气候异常变化加剧,渍涝等灾害将更加频繁发生,近年我国的洪涝灾害也有面积增大、危害加重趋势。
植物的呼吸作用是其新陈代谢的重要途径之一,其途径和强弱直接影响到生长发育。渍涝灾害发生后,过多的土壤水分必然导致氧气含量骤然减少,最直接的伤害是造成植株根际环境缺氧,根系的呼吸作用、营养物质包括水分吸收严重受阻,继而引起地上部分损伤甚至死亡,最终影响产量和品质[5-10]。花生为我国重要的油料作物之一,湿涝是我国南方地区花生芽苗期频发的灾害。目前花生水分胁迫的研究以干旱居多,有关渍涝下花生种子发芽期呼吸生理的研究鲜见报道。本研究拟采用耐渍涝能力不同的3个品种,测定渍涝胁迫下几种关键性呼吸酶活性的动态变化,试图初步探明不同耐渍涝性品种的呼吸途径差异及机理。
1 材料与方法
1.1 试验材料
选取3个经过验证[10]在大田条件下,具有耐涝性差异的花生栽培品种(ArachishypogaeaL.)豫花15(耐涝)、中花8号(较耐涝)、中花4号(不耐涝)为供试品种。
1.2 试验设计
精选后的种子用10%过氧化氢消毒15 min后,蒸馏水漂洗4次。将种子放在具2层湿滤纸的无菌培养皿中(直径13 cm),每皿中均匀分散摆放25粒种子,加入蒸馏水,要求水深达到皿口最高处,以便浸没种子。为避免种子浮在水面,均用2层消毒的透气纱布覆盖在水面上。培养皿不加盖、25℃培养箱中暗培养。
设置浸泡时间分别为6(对照)、12、24、48、72、96、120、144、168 h后排除水分,移到湿润培养皿中正常发芽,每24 h加水1次,加水程度以保持培养皿中滤纸湿润即可,重复3次,每24 h观测记录一次,并选取已发芽的种子(当胚根长达到3 mm长度时,设定为正常发芽的记载标准)进行测定。
1.3 测定项目与方法
琥珀酸脱氢酶(SDH)活性和乳酸脱氢酶(LDH)活性的测定分别采用SDH和LDH试剂盒测定,均由南京建成生物工程研究所提供。乙醇脱氢酶(ADH)活性的测定采用紫外分光光度计法(汤章城等[11])。
2 结果与分析
2.1 淹水条件对不同品种发芽期有氧呼吸途径主要酶活性的影响
图1可见,在种子发芽阶段持续淹水胁迫下,3个品种的SDH活性均呈现从高向低的起伏状下降趋势,其中,豫花15号的SDH活性最先变化,在12 h有最大峰值,之后呈下降趋势,在72 h有所回升,到96 h达第二峰值, 144 h出现第三峰值。中花8号在6~72 h内都为活性上升趋势,并在72 h达最大峰值,之后逐步下降,在168h回升,是其观测周期内的第二峰值。中花4号在24~48 h内活性骤增,48 h达到其最大峰值,之后呈逐渐下降趋势直至观测结束。三个品种的SDH最大活性值以豫花15号最高。
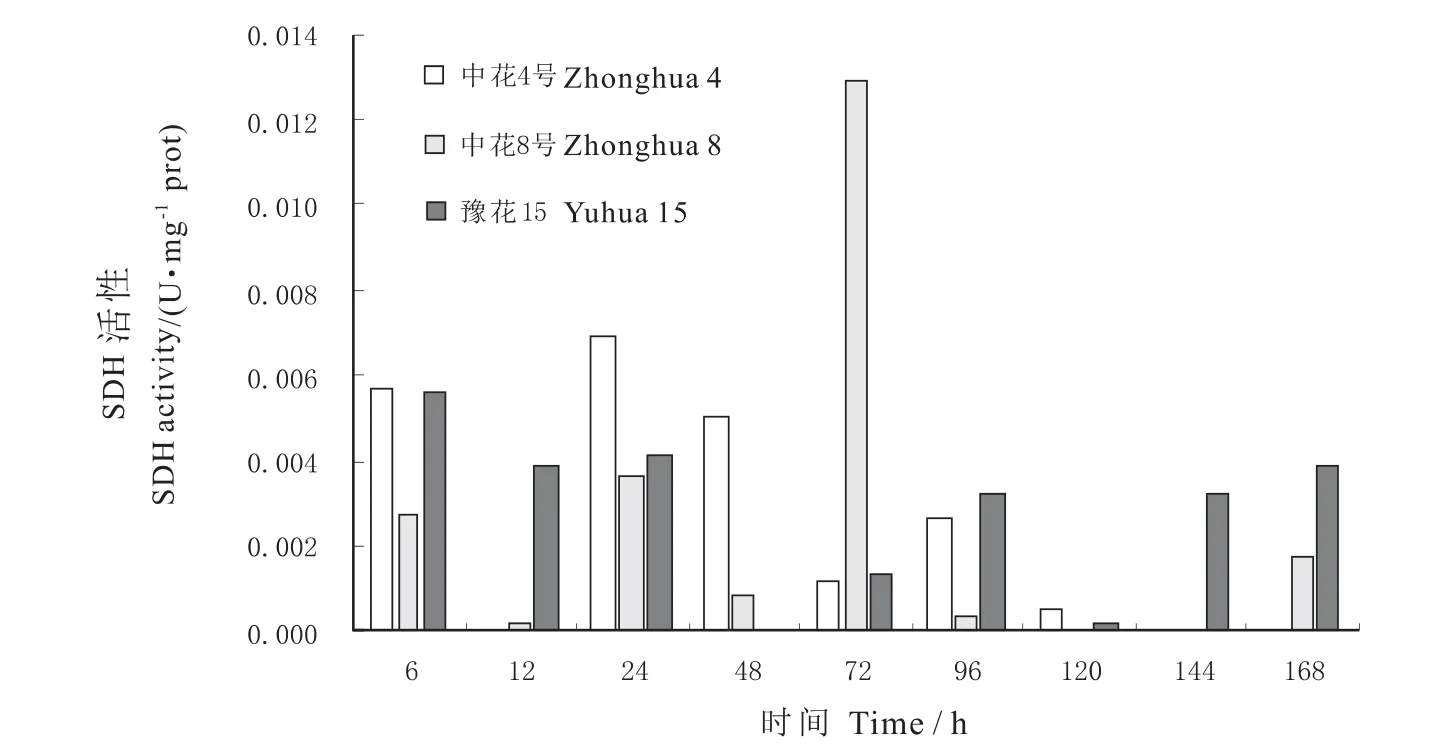
图1 不同花生品种发芽期琥珀酸脱氢酶(SDH)比活性变化Fig.1 The changes of SDH activity in seed germination stage of different peanut varieties under waterlogging tolerance condition
2.2 发芽期淹水条件下不同品种厌氧呼吸呼吸途径酶活性差异
2.2.1 乳酸脱氢酶(LDH)活性
图2可见,在种子发芽阶段持续淹水胁迫下,3个品种的LDH活性呈中高、前后均低的起伏状变化趋势,说明短期(6~48 h)淹水不足以诱导LDH活性,而过度淹水(96~168 h)则抑制LDH活性。其中,中花4号的LDH活性在6~72 h呈上升趋势,72~96 h内转为下降,96 h之后又有所回升。中花8号的LDH活性在72 h处达最大峰值,此前的12 h、此后的120 h出现2个小高峰。豫花15号的LDH活性变化趋势与中花4号相似。3个品种的LDH活性最大峰值相比,以豫花15号最高。
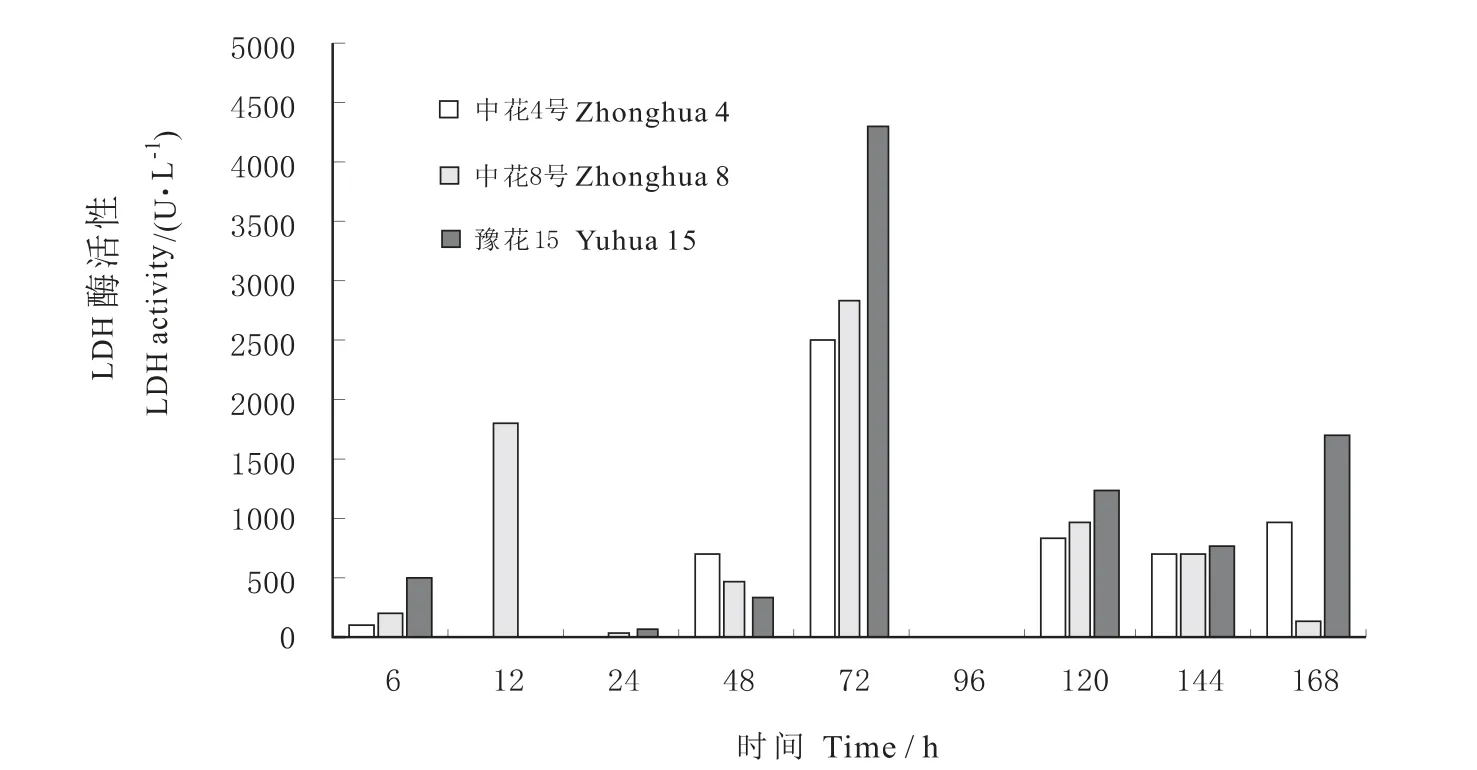
图2 不同花生品种发芽期乳酸脱氢酶(LDH)活性变化Fig.2 The changes of LDH activity in seed germination stage of different peanut varieties under waterlogging tolerance condition
2.2.2 乙醇脱氢酶(ADH)活性
图3可见,种子发芽阶段持续淹水胁迫下,3个品种的ADH活性均呈现与SDH截然不同、与LDH相似的单峰状变化趋势,最高峰均出现在72 h,峰值以中花8号最高,豫花15号最低,仅为前二者的1/3,说明耐涝品种的ADH活性不易被缺氧所诱导,而敏感品种能被快速诱导。其中,中花4号的ADH活性在发芽期淹水阶段的前期(12 h)、中期(72 h)、后期(96 h)均最高,96 h之后活性极低至无;中花8号的ADH在48 h、72 h处急剧上升达到最大值,此前此后均极低;豫花15号的ADH活性也集中出现在24~96 h,此前此后均极低。
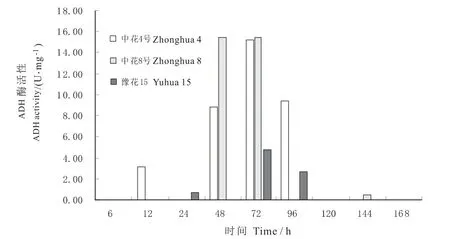
图3 不同花生品种发芽期乙醇脱氢酶(ADH)活性变化Fig.3 The changes of ADH activity in seed germination stage of different peanut varieties under waterlogging tolerance condition
3 讨 论
植物的呼吸作用分为有氧、无氧呼吸两种。在正常供氧情况下,植物多进行有氧呼吸,在有氧呼吸途径中葡萄糖被氧化生成丙酮酸,然后进入三羧酸循环(TAC),被完全氧化生成CO2和H2O,并释放能量,琥珀酸脱氢酶(SDH)是TAC循环的限速酶,其活性的高低直接影响到三羧酸循环的运行效率。而当植物处于缺氧逆境胁迫条件下,植物体为了维持短期的生活机能,将会进行无氧呼吸来产生能量,此时葡萄糖生成的丙酮酸在TAC循环中不会被完全氧化,并低效、少量地释放能量。丙酮酸在乳酸脱氢酶的催化作用下,被途径中生成的NADH还原为乳酸;或者在丙酮酸脱羧酶作用下脱羧生成乙醛,再在乙醇脱氢酶的作用下转变成乙醇。若长期进行无氧呼吸,将使得乳酸和乙醇在植株体内大量积累,最终将造成植株中毒死亡。
本试验发现,3个花生品种正常供水时SDH活性高于湿涝胁迫,但LDH、ADH活性很低。在种子发芽阶段6~168 h淹水胁迫下,3个品种的SDH活性均呈现从高向低的起伏状下降趋势,与前人关于淹水玉米根系SDH活性下降的结论相似[12];LDH活性能被缺氧胁迫迅速诱导,呈中高、前后低的起伏状变化趋势,说明短期(6~48h)淹水不足以诱导LDH活性规律变化,而过度淹水(96~168 h)则抑制LDH活性,这与喜树、黄瓜幼苗受涝渍胁迫后大幅升高的情形相似[17-18];ADH活性也能被缺氧胁迫迅速诱导,呈单峰状变化趋势,与前人在其他植物中得出的ADH活性持续上升的结论总体相似[14-18],但与彭克勤等[22]的水稻ADH先降低后升高的结论有所不同,这可能与植物所处生长阶段、淹涝时间长短密切相关,与植物种类不同也有一定联系。3个品种相比较,豫花15号的SDH、LDH活性峰值最大,ADH活性峰值最低;中花8号的SDH活性峰值最低、LDH活性峰值居中、ADH活性峰值最高,中花4号的SDH活性峰值较高、LDH活性峰值最低、ADH活性峰值高。说明耐涝品种的有氧呼吸活性强,乳酸发酵强度也高,乙醇发酵强度弱,而敏感品种的乙醇发酵活性易被诱导升高,这也说明不同耐涝性品种呼吸途径的复杂性。这一结果与前人研究结论一致[19-22]。
不同氧气条件下,植物呼吸途径的改变是其适应环境的重要机制[23-26]。本试验中,根据SDH、LDH和ADH三者活性随淹涝时间的变化趋势,推测在不同供氧条件下的一种互动关系及可能机制,在淹涝初期,由于花生种子体内还有留存的氧气,TAC循环在短期内尚可维持将丙酮酸完全氧化的状态,所以此时的有氧呼吸活性大于无氧呼吸,SDH活性较高,而LDH和ADH的活性尚未得到大幅诱导启动,维持在较低水平上;在淹涝中期,花生种子体内残留的氧气基本上已被消耗殆尽,种子为了维持代谢生长,无氧呼吸逐渐取代有氧呼吸,SDH活性持续降低,LDH和ADH活性迅速上升。但是由于无氧呼吸同样会产生丙酮酸,所以当有丙酮酸产生时,三羧酸循环的运行水平高,SDH的活性高,当丙酮酸含量低时,三羧酸循环的运行水平低,SDH的活性低。
致谢:感谢湖南农业大学作物生理与分子生物学省部共建重点实验室屠乃美教授、李海林老师、刘红梅老师、周双鹏与孙媛同学在分析测试工作中给予的帮助。
猜你喜欢
少先队活动(2022年9期)2022-11-23
成都信息工程大学学报(2022年4期)2022-11-18
山西医科大学学报(2022年6期)2022-08-04
中国临床医学影像杂志(2022年6期)2022-07-26
牡丹江医学院学报(2021年5期)2021-12-05
家庭科学·新健康(2021年5期)2021-06-21
科学大众(2021年10期)2021-05-20
岭南急诊医学杂志(2020年6期)2021-01-14
食品与生物技术学报(2020年3期)2020-05-19
家庭医药·快乐养生(2017年11期)2017-11-21
