
湖南省棉花黄萎病菌致病力分化及致病型分布研究
2019-04-10 08:28李敏李彩红赵瑞元刘冰蕾张志刚
棉花学报 2019年2期
李敏,李彩红,赵瑞元,刘冰蕾,张志刚*
(1.湖南农业大学,长沙410128;2.湖南省棉花科学研究所,湖南常德415101)
棉花黄萎病(Verticillium wilt)是1 种世界性的土传真菌病害,对棉花生产造成了严重的威胁。该病自20 世纪30年代由美国传入中国,历经几十年时间已蔓延到全国各个棉区,并造成严重的经济损失,是棉花可持续生产的主要障碍之一[1-4]。在中国,引起黄萎病发生的主要病原物是大丽轮枝菌(Verticillium dahliae)[5]。大丽轮枝菌致病力变异性强,在与寄主协同进化及受到生态环境变化影响的过程中常分化出新致病力类型[1],而不同致病力的大丽轮枝菌在同一寄主体内诱发的症状也会有所差异。因此,不断研究其致病力分化对于抗黄萎病品种选育及棉花生产的可持续发展具有重要意义。
长期以来,很多学者对中国各主要植棉区棉花黄萎病菌进行了致病力分化研究[1]。湖南省棉花黄萎病的发生呈日益加剧的态势,但到目前为止,有关湖南省棉区棉花黄萎病菌情况尚无系统性研究,这在一定程度上间接阻滞了湖南省抗病育种进程。本研究以湖南省各主产棉县(区)分离的棉花黄萎病菌为研究对象,分析其生理分化和致病力、落叶型分布情况,为该地区棉花抗病品种选育推广及棉花黄萎病综合防控提供依据。
1 材料与方法
1.1 供试菌株
2013年、2014年在棉花黄萎病发病高峰期(7―8 月)从湖南省各主产棉县(区)采集发病典型的黄萎病病株,采用常规组织分离法和稀释平板法[5],共获得77 个菌株单孢,其中鼎城区14株,南县26 株,澧县10 株,汉寿县26 株,安乡县1 株。所有菌株于4 ℃低温冰箱保存。
1.2 培养特性观察
将保存的各黄萎病菌菌株单孢转接至马铃薯葡萄糖琼脂培养基(Potato dextrose agar medium,PDA 平板)上,于25 ℃黑暗培养15 d,观察记载各菌株的菌落形态、菌丝生长及菌核产生情况,根据培养特性,将供试病菌划分为菌核型、菌丝型和中间型[6]。
1.3 生长速率测定
选择31 个菌株,分别取5 μL 含量106mL-1的孢子悬浮液,滴入查氏(Cazpek)培养基的中央,放置在25 ℃培养,每个菌株重复4 次。采用十字交叉法,于第5、7、9、11、13 天测量菌落直径。
1.4 产孢量测定
将用于生长速率测定的31 个菌株在PDA平板上25 ℃恒温培养5 d,沿菌落边缘取直径5 mm 的菌丝块置于Cazpek 培养基(40 mL),25 ℃,150 r·min-1振荡培养6 d 后,以血球计数板(每个菌数40 个小格)测定其产孢量。
1.5 致病力测定
选择黄萎病抗性不同的6 个棉花品种作为鉴别寄主,包括抗病品种鲁棉研28、中棉所41、豫棉2067,耐病品种中棉所35,感病品种中棉所8 号和冀棉11 号。
将硫酸脱绒的鉴别寄主种子浸泡12 h 后沥干水分,在温室条件下播种到营养纸钵中,每重复6 个钵,待棉苗出土后每钵留苗3~4 株,共3次重复。
将上述31 个供试菌株活化后接种到Cazpek培养基中,25 ℃黑暗振荡(150 r·min-1)培养7 d左右,用4 层纱布过滤制成孢子悬浮液,在显微镜下用血球计数板计数,将孢子含量调到1.0×107mL-1,现用现配。待棉苗长到1 片真叶平展时,参照朱荷琴等[7]蛭石沙土无底纸钵定量蘸菌液法进行接种,接菌量为每钵15 mL,以接等量清水作为对照。接种后15 d 开始调查,按全国统一病情分级标准进行分级调查,记录病情。以接种后第24 天调查的结果计算病情指数(Disease index,DI),并以DI 作为划分反应类型(Reaction type,RT)的标准:免疫型(I),病情指数为0.0;高抗型(HR),病情指数为0.1~10.0;抗病型(R),病情指数为10.1~20.0;耐病型(T),病情指数为20.1~35.0;感病型(S),病情指数为35.0 以上[8]。
1.6 落叶型检测
选择24 个供试菌株,采用CTAB 法提取DNA,以大丽轮枝菌落叶型菌株特异性引物D-1/D-2[9](5′-CATGTTGCTCTGTTGACTGG-3′/5′-GACACGGTATCTTTGCTGAA-3′) 和非落叶型菌株特异引物ND-1/ND-2[9](5′-CAGGGGATACTGGTACGAGACG-3′/5′-ATGAGTATTGCCGATAAGAACA-3′) 进行聚合酶链式反应(Polymerase chain reaction,PCR)扩增。引物由苏州金维智生物科技有限公司合成。
PCR 反应体系:2×TaqPCR MasterMixⅠ2.5 μL(天根生化科技有限公司)、引物各1.0 μL、模板DNA 0.5 μL、ddH2O 补足至25 μL。反应程序:94 ℃预变性3 min;94 ℃变性50 s;退火(温度:D引物41 ℃,ND 引物50 ℃)50 s;72 ℃延伸1 min;共循环35 次;72 ℃延伸10 min;4 ℃保存。在质量分数1.0%的琼脂糖凝胶上电泳检测。
1.7 统计分析
以SAS 9.1.3 统计分析软件中的聚类分析程序对31 个棉花黄萎病菌菌株的致病力测定结果进行聚类分析;以SPSS 软件对致病力、生长速率及产孢量进行相关性分析。
2 结果与分析
2.1 供试菌株的培养特性
供试菌株在PDA 平板上的菌落形态差异明显,根据微菌核产量及菌落特性,将湖南棉花黄萎病菌划分为菌核型、菌丝型和中间型3 种培养类型,且每种类型的各个菌株形态多样(图1)。
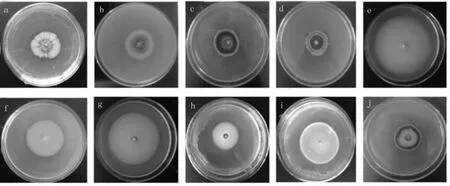
图1 不同类型供试菌株培养特性Fig.1 Culture characteristics of different type strains
菌核型菌株共11 个,占14.28%(表1)。该类型表现为产生大量黑色微菌核,中央部分气生菌丝发达。根据其形态可分为4 种:(1)黑色菌核部分呈放射状,菌丝致密;(2)整个菌落表面气生菌丝发达,外围形成规则的白色菌丝圈;(3)菌落边缘呈现不规则白色菌丝圈;(4)菌落边缘不整齐,内部形成白色菌丝圈。
菌丝型菌株共33 个,占42.85%。根据其形态可分为5 种:(1)无菌核产生,菌丝疏松,有轮纹;(2)无菌核产生,菌丝致密,表面有少量气生菌丝;(3)菌落中央有少量黑色菌核,菌丝致密,有轮纹;(4)菌落中央有少量黑色菌核,无轮纹产生;(5)菌落边缘呈现白色环状圈。
中间型菌株共33 个,占42.85%。此类菌株的菌核大多分布在菌落周围,形成黑色圆环,菌落中间为白色。
2.2 供试菌株生长速率和产孢量
对31 个菌株进行生长速率和产孢量测定,结果(表2) 表明:供试菌株生长速率为1.22~2.54 mm·d-1,分离自澧县的XVd60 及汉寿的XVd61 菌株生长最快,分离自南县的XVd40 生长最慢;菌株之间产孢量差异较大,其中XVd14产孢量最低,为5.3×106mL-1,XVd2 产孢量最高,为40.6×106mL-1。
2.3 供试菌株致病力鉴定
根据平均病情指数以最小距离法对供试菌株的致病力进行聚类,31 个黄萎病菌菌株被划分为致病力强的Ⅰ型、致病力弱的Ⅱ型和致病力中等的Ⅲ型(图2)。其中:Ⅰ型菌株仅有分离自汉寿的XVd78,占所有供试菌株的3.2%(表3),对6个鉴别寄主的平均病情指数为43.8,抗病品种中棉所41 和豫棉2067 也表现为感病,病情指数分别为38.7 和39.8。Ⅱ型菌株在6 个鉴别寄主的平均病情指数为23.6~26.5,占12.9%,分布在澧县、南县和汉寿。Ⅲ型菌株在6 个鉴别寄主的平均病情指数为29.1~41.9,占比最大,为83.9%,
在湖南省各个地区均有分布。综合31 个供试菌株对所有鉴别寄主的病情指数分析,抗病水平最高的品种是鲁棉研28,其次是中棉所41,抗病水平最低的为冀棉11 号。
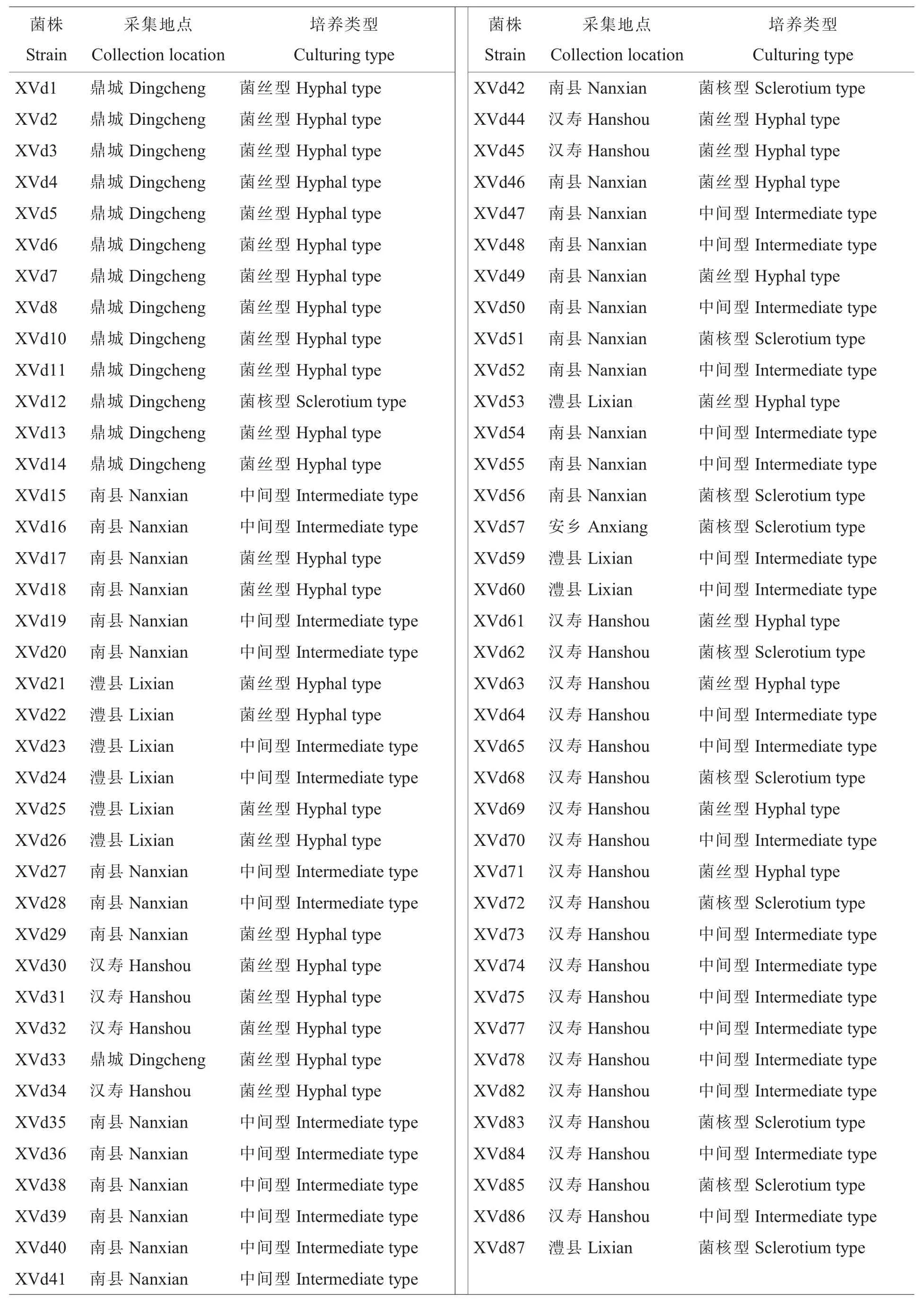
表1 77个菌株培养类型Table 1 Culturing types of 77 strains
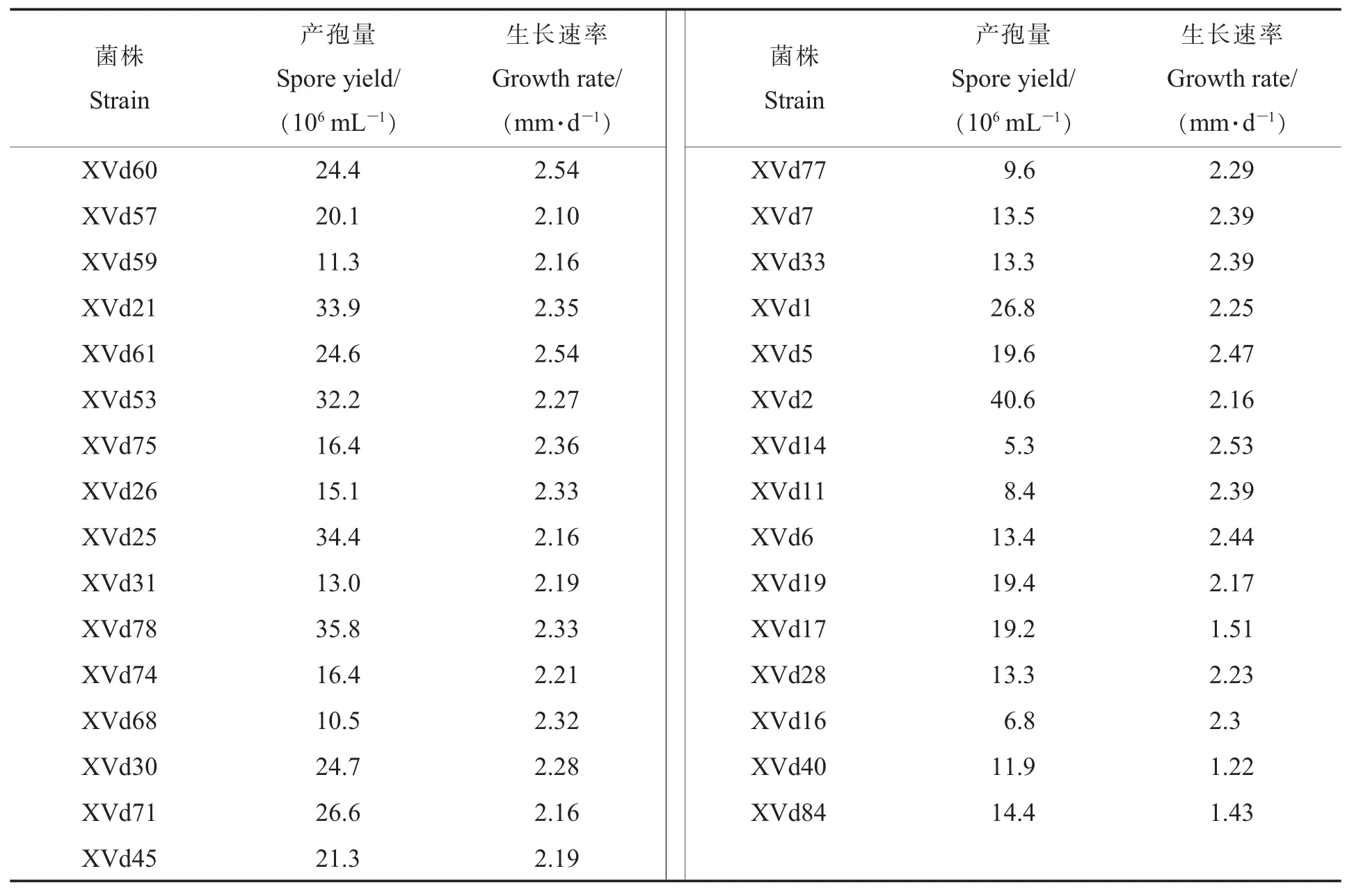
表2 31个菌株产孢量及生长速率Table 2 The sporulation quantity and growth rates of 31 strains

图2 31个菌株基于平均病情指数的系统聚类图Fig.2 Dendrogram of 31 strains based on averaged disease indexes
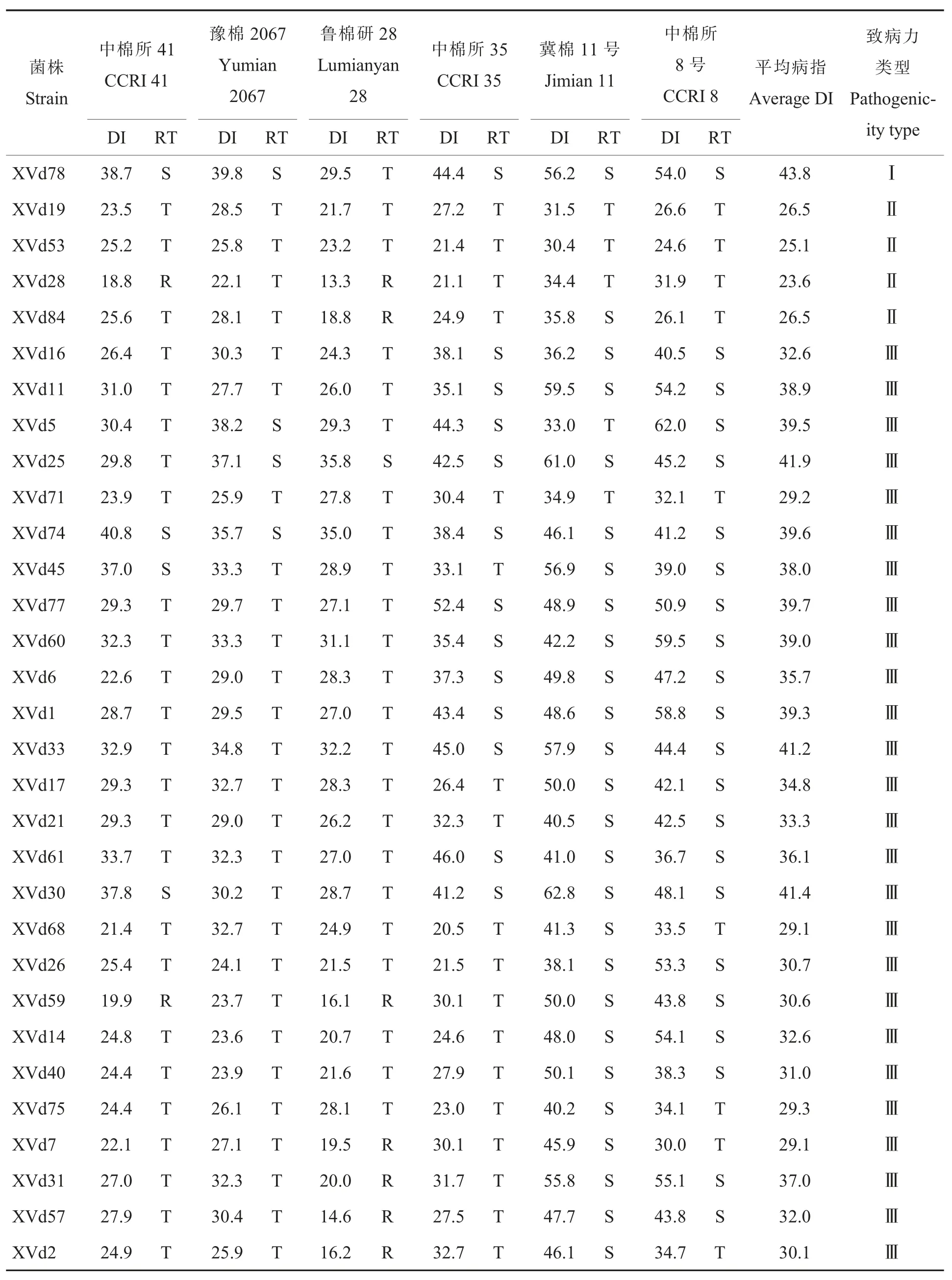
表3 31个棉花黄萎病菌株对6个鉴别寄主的致病性Table 3 Pathogenicity of 31 strains of V.dahliae to six different hosts
2.4 供试菌株致病型PCR检测
如图3 所示,24 个供试菌株由落叶型特异性引物均扩增出大小为550 bp 的目的条带,而由非落叶型特异性引物则扩增不出条带。表明湖南地区棉田采集到的黄萎病菌主要为落叶型菌株。

图3 D-1/D-2 PCR检测供试菌株的致病型Fig.3 The pathotype of the tested strains by PCR assay with primers D-1/D-2
3 讨论
3.1 湖南省棉花黄萎病菌的培养类型
77 个菌株可按照培养特性被分为菌核型、菌丝型及中间型,其中菌核型占比最小,菌丝型和中间型占比相等,表明湖南地区的棉花黄萎病菌以菌丝型和中间型为主。此结果与李彩红等[10]于2015年对湖南常德地区棉花黄萎病监测结果有所差异,后者中菌丝型占比最大,达67.6%。原因可能是湖南地区棉花黄萎病菌分化速度较快,且优势种群正处于变化中。而在全国范围,2007年以后江苏[8]、河南[11]、新疆[12]、河北[13]等地报道的棉花黄萎病菌以菌核型为主,表明菌核型正在这些地区成为优势种群,这一趋势与湖南省情况不同。本研究中菌核型和菌丝型2 种类型的菌株菌落表现出形态多样性,且跟其他省份所报道的形态差异较大[8,11-12,14-15],这表明棉花黄萎病菌在不同地区的分化差异较大。
3.2 湖南省棉花黄萎病菌致病力分化情况
本研究在鉴别寄主的选择上参考了袁媛等[16]的研究,该研究以我国三大棉区黄萎菌菌株为对象,筛选确定了一套棉花黄萎病菌致病力测定的鉴别寄主,这在一定程度上增强了本研究致病力测定结果的准确性。从致病力测定结果来看,湖南省各主产区棉花黄萎病菌存在较大差异,且以III 型为主,占供试菌株的83.9%。所有6 个鉴别寄主对77.4%的供试菌株表现耐病或感病,表明湖南省棉花黄萎病菌具有较强的致病力。31 个供试菌株致病力与生长速率之间的相关系数为0.190,与产孢量之间的相关系数为0.155,表明致病力与生长速率、产孢量均无显著相关性,这与林玲等[8]、刘廷利等[12]研究结果一致,也在一定程度上说明湖南棉花主产区黄萎病菌分化严重。
棉花黄萎病菌致病力与培养特性的相关性,一直是学者们研究的热点。朱荷琴等[17]和李艳等[18]研究认为,棉花黄萎病菌株致病力与生长速率呈正相关,且与培养特性存在相关性;林玲等[19]研究报道,江苏省棉花黄萎病菌致病力与产孢量的关系不明显。近年来,多数研究棉花黄萎病致病机制的报道则表明,病原菌的生长速率、产孢量与致病力有一定相关性[20-22];但病原菌对寄主致病并不是单一的信号传导事件,往往是多种信号途径交叉影响的结果[23-24]。
3.3 湖南省棉花黄萎病菌致病型
本研究利用PCR 技术检测湖南省棉花主产区黄萎病菌的致病型,落叶型菌株占100%。该结果从分子水平上论证了落叶型菌株是当前湖南省棉花黄萎病菌优势种群,与王彦等[25]、徐飞[26]的研究结果“华中地区棉花主产区以落叶型黄萎病菌为主”一致。由于本试验仅选取了24 个菌株为研究对象,尚需扩大供试菌株样本量进行进一步研究。此外,大丽轮枝菌特异性引物PCR 法是被普遍认同的鉴定棉花黄萎病致病类型的方法,但朱荷琴等[24]认为能否侵染棉花并引发典型的落叶症状才是判断落叶型菌株最重要的指标。因此,落叶型菌株是否为湖南省优势种群仍需扩大样本量并结合生物测定进行进一步探究。
4 结论
本研究对湖南省棉花主产区分离到的77 个棉花黄萎病菌菌株进行了培养特性分析,进一步分析了其中31 个菌株的生长速率、 产孢量及致病力,明确了湖南省棉花黄萎病菌主要培养类型由菌丝型变化为菌丝型和中间型,致病型以落叶型为主,致病力以Ⅲ型为主。同时,湖南省棉花黄萎病菌分化速度较快且致病力分化严重,针对这一现状结合简单重复序列间扩增(Inter-simple sequence repeat-PCR,ISSR-PCR) 等技术手段对其进行持续监测是十分必要的。
猜你喜欢
中国生物防治学报(2022年4期)2022-09-16
作物学报(2022年9期)2022-07-18
陕西农业科学(2022年2期)2022-03-28
农业科技与信息(2021年4期)2021-12-05
新农村(浙江)(2021年2期)2021-11-30
西北农业学报(2021年5期)2021-06-28
食用菌(2021年3期)2021-06-03
浙江农业学报(2021年5期)2021-05-31
中国食用菌(2021年1期)2021-03-12
中国棉花(2021年11期)2021-03-05
