
拮抗内源性孤啡肽可通过下调microRNA-1途径抑制大鼠再灌注性心律失常
2019-04-01 11:20:44任晓芬王一迪李占峰
中国药理学通报 2019年4期
熊 畅,任晓芬,韩 毅,2,王一迪,李占峰
(1. 山西医科大学麻醉学系,山西 太原 030000;2. 山西医科大学第二医院麻醉科,山西 太原 030001)
目前,临床治疗急性心肌梗死患者最有效的方式是恢复梗死区血运,然而,在血流再通后可出现缺血/再灌注损伤(ischemia/reperfusion injury, IRI)。再灌注性心律失常(reperfusion arrhythmia, RA)是心肌IRI最重要的临床表现,是目前临床判断血流恢复的指标之一,但同时恶性RA也是导致患者预后不良,甚至死亡的重要原因[1-2]。
microRNAs是非编码微小RNA,能够从转录后水平调节心肌离子通道及传导结构的表达。研究发现,microRNAs对于心肌细胞钠、钾、钙等多种离子通道,以及心肌传导结构如缝隙连接等均能发挥调节作用。病理条件下,异常表达的各种微小RNA通过调控离子通道以及传导结构,引起心肌电重构以及结构重构,直接导致各种心律失常的产生[3-4]。microRNA-1(miR-1)是目前发现的一种肌肉特异性微小RNA,已有实验证明心肌缺血状态下升高的miR-1能够抑制GJA1(编码connexin 43, Cx43)以及KCNJ2(编码内向整流钾通道的核心亚基kir2.1)基因表达,导致心律失常的发生[5]。
孤啡肽(nociceptin/orphanin FQ, N/OFQ)是一种具有神经递质功能的内源性阿片肽,本课题组前期研究发现,急性心肌缺血后,心肌组织N/OFQ表达明显增多,并且拮抗内源性N/OFQ能够发挥抗缺血性心律失常作用[6],但其是否参与RA的发生尚不清楚。本研究拟观察内源性N/OFQ对RA、miR-1及其下游靶点的影响,探讨内源性N/OFQ对RA的作用及其调节机制。
1 材料与方法
1.1材料
1.1.1实验动物 SPF级健康成年SD大鼠24只,♂,体质量(260±20)g,购自中国人民解放军军事科学研究院实验动物中心,许可证号:SCXK(军)2012-0004,适应性喂养1周。动物实验已通过山西医科大学伦理委员会批准。
1.1.2试剂 UFP-101(Tocris Cookson公司);TRIzol(TaKaRa公司);cDNA合成试剂盒(Thermo Scientific公司);SYBR Green qPCR Master Mix(2×)(Fermentas公司);抗Cx43、kir2.1、GAPDH抗体(Abcam公司)。
1.1.3仪器 AlC-V8小动物呼吸机(上海澳尔科特公司);RM6240BD电生理信号记录仪(成都仪器厂);MX3005P实时荧光定量PCR仪(上海拜力生物科技有限公司);ChemiDocXRS凝胶成像分析系统(Bio-Rad公司)。
1.2动物模型的建立及分组用随机数字表法分为3组(n=8):假手术组(Sham组)、缺血/再灌注组(I/R组)和孤啡肽受体拮抗剂UFP-101预处理组(U+I/R组)。Sham组仅开胸穿线,但不结扎冠脉;I/R组结扎左冠状动脉前降支缺血30 min,随后再灌注120 min;U+I/R组在再灌注前10 min经尾静脉按1 μL·g-1注射特异性孤啡肽受体拮抗剂UFP-101(10-9mol·L-1),其余2组给予等容量生理盐水。大鼠称重后,以质量分数为25%的乌拉坦(1.25 g·kg-1)进行腹腔注射麻醉,将大鼠仰卧位固定于操作台上,连接生物信号采集系统,记录标准Ⅱ导联心电图。气管切开置入气管导管,连接小动物呼吸机施行机械通气,呼吸机潮气量设为8 μL·g-1,呼吸频率70次/min。稳定15 min后,在左侧第4肋间隙开胸,以5-0无损伤缝线的弯针从左心耳右缘进针,肺动脉圆锥左缘出针以套管结扎左冠状动脉前降支。而后可见Ⅱ导联心电图出现ST段逐渐抬高,并渐与QRS波融合,心肌前壁组织颜变苍白色随后紫绀,可有心律失常出现,提示造模成功。缺血30 min后,松开结扎线使血管再通(松开套管前10 min经尾静脉给予UFP-101或等量生理盐水),心外膜出现充血,同时心电图ST段有所回落,心肌呈再灌注状态,再灌注120 min后处死大鼠,取缺血区心肌。
1.3心律失常统计及评分对心律失常进行评定(Fig 1),记录再灌注后室性早搏(ventricular ectopic beats, VEB)的发生个数(包含单发室早、室早二联律和三联律)、室速(ventricular tachycardia, VT)和室颤(ventricular fibrillation, VF)的发生次数以及持续时间,并根据参考文献[7]对大鼠心律失常进行评分。评分标准如下,0分,共出现<50个VEB(包括室早二联律、室早三联律);1分,出现≥50个VEB;2分,出现1~5次VT;3分,出现≥ 6次VT;4分,出现1次可逆性VF;5分,出现2~5次可逆性VF;6分,出现不可逆性VF。

Fig 1 Examples of arrhythmias
A: Normal ECG; B: VEB; C: Bigeminy; D: Trigeminy; E: ST elevation; F: VT; G: VF.
1.4qRT-PCR检测将液氮保存的心肌组织加入DEPC水处理过的研钵中,持续加入液氮研磨至粉末。称取20~30 mg心肌,利用TRIzol提取总RNA,然后逆转录合成cDNA,再按SYBR Green qPCR Master Mix(2×)试剂盒步骤进行扩增。引物由TaKaRa生物有限公司合成(Tab 1)。miR-1采用U6作为内参基因,Cx43、kir2.1 mRNA采用GAPDH基因作为内参。反应条件,预变性:95 ℃ ,60 s;扩增(40个循环):95 ℃,20 s;57 ℃,30 s;72 ℃,60 s;熔解曲线:95 ℃,30 s;60 ℃,60 s;95 ℃,30 s。在每个循环延伸末收集荧光信号,绘制扩增曲线,检测各样本的域值循环数(CT值),以2-△△CT法对目的基因进行相对定量分析。

Tab 1 Primer sequence of qRT-PCR
1.5Westernblot检测将液氮中保存的心肌切成小块,将部分组织按10 mg加100 μL蛋白裂解液,冰上孵育并振荡充分裂解,4 ℃、12 000×g离心30 min,取上清液,分装-80 ℃保存。分装蛋白利用BCA法进行蛋白定量,依次进行SDS-PAGE凝胶电泳、转膜、5%脱脂奶粉封闭2 h后,分别加入一抗Cx43(1 ∶2 000)、kir2.1(1 ∶1 000)、GAPDH(1 ∶1 500)摇匀后,4 ℃孵育,摇床过夜,加入二抗HRP-IgG(1 ∶2 500),室温孵育1 h 。TBST清洗,3次×10 min。将PVDF膜条带表面滴加ECL显影液,放入凝胶成像仪暗室中,应用Quantity One图像分析软件测定各蛋白条带的灰度值,以GAPDH为内参反映其相对表达水平。
2 结果
2.1动物模型建立情况实验过程出现心律失常只做记录,未进行抗心律失常处理。共27只大鼠参与实验过程,其中因实验操作不当或结果有误弃除3只,剩余24只大鼠均存活至实验结束。
2.2心律失常发生情况如Tab 2所示,与Sham组比较,I/R组以及U+I/R组均出现明显的再灌注性心律失常(P<0.01)。与I/R组比较,U+I/R组再灌注期间室早次数、室速和室颤发生次数、持续时间及致心律失常评分明显下降(P<0.01),而室速和室颤的发生率稍有减少,但差异无统计学意义。
2.3拮抗内源性N/OFQ对心肌miR-1表达的影响如Fig 2所示,与Sham组相比,I/R组心肌miR-1表达明显增多(P<0.01);与I/R组相比,U+I/R组miR-1表达明显下降(P<0.05)。

Fig 2 miR-1 expression at 120 min after n=8)
*P<0.05,**P<0.01vssham;#P<0.05vsU+I/R
2.4拮抗内源性N/OFQ对心肌Cx43、Cx43mRNA以及kir2.1、kir2.1mRNA的影响与Sham组相比,I/R组Cx43、Cx43 mRNA以及kir2.1表达降低(P<0.05),kir2.1 mRNA降低无统计学意义。与I/R组相比,U+I/R组Cx43、kir2.1表达明显增加(P<0.05),Cx43 mRNA及kir2.1 mRNA表达虽有增多,但差异无统计学意义(Fig 3、4)。
3 讨论
RA是血运重建后发生的二次损伤,影响患者预后,甚至导致死亡,多种因素参与其病理生理过程。再灌注早期,心肌表现为氧自由基增多、钙超载、炎症因子聚集等,与此同时,心脏自主神经以及感觉神经被激活,释放多种神经递质/调质,共同参与再灌注后心脏的损伤与保护,其中N/OFQ即是心脏感觉神经末梢释放的神经肽之一[8-9]。
N/OFQ及其受体广泛分布于机体多种组织器官,包括心血管系统、神经系统以及消化系统等,参与疼痛、抑郁、免疫等多种病理生理过程[10]。孤啡肽受体(NOP/ORL1)与经典阿片受体存在高度同源性,但其功能与三大经典阿片受体(μ、δ、κ)却截然不同,故又称为孤儿阿片受体(ORL1)[11]。研究发现,N/OFQ对神经细胞多种离子通道均能发挥抑制作用[12],本课题组前期研究亦发现,N/OFQ能够调节大鼠心肌细胞动作电位时程,参与缺血性心律失常过程[6]。而缺血/再灌注状态下,N/OFQ能否影响RA的发生尚无报道。本研究结果表明,拮抗内源性N/OFQ同样可降低再灌注后VEB、VT、VF等快速性心律失常的发生。
Tab 2 Comparison of cumulative reperfusion-induced arrhythmias in
**P<0.01vssham;##P<0.01vsU+I/R

Fig 3 Expression of kir2.1 and Cx43
*P<0.05,**P<0.01vssham;#P<0.05vsU+I/R
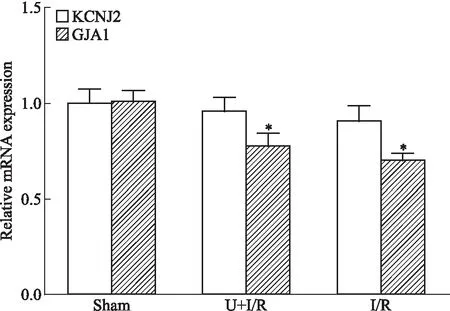
Fig 4 Expression of KCNJ2 and GJA1
*P<0.05vssham
既往研究表明,心肌缺血状态下,miR-1表达增多能够抑制Cx43及kir2.1蛋白表达,引起电生理紊乱[4-5]。Cx43是心肌细胞间电信号传导的重要连接蛋白,是心肌同步化收缩的结构基础,而kir2.1蛋白则是内向整流钾通道(IK1)的核心亚基,对于心肌细胞静息电位的维持有着重要影响。然而,再灌注期间,miR-1的表达是增多还是降低,尚有不同结果报道[13-14]。本研究通过制备大鼠缺血/再灌注模型,结果显示,再灌注120 min后,心肌miR-1表达明显增多,并发现其下游靶点Cx43、kir2.1表达出现转录后抑制表现。由此推测,再灌注后miR-1的增多可导致Cx43及kir2.1表达抑制,进而引起RA,本结果与文献报道一致[14]。
在明确N/OFQ以及miR-1均是RA的影响因素后,本研究根据前期研究结果,选择特异性孤啡肽受体拮抗剂UFP-101最佳剂量(10-9mol·L-1, 1 μL·g-1)进行尾静脉注射给药[6]。结果显示,拮抗内源性N/OFQ后,miR-1的表达水平明显下降,并且Cx43及kir2.1表达有所上调。结果提示内源性N/OFQ是导致再灌注期间miR-1上调的原因之一,而N/OFQ对于RA的调控很可能是通过上调miR-1,抑制Cx43及kir2.1表达而实现。值得注意的是,本实验通过比较Cx43 mRNA及kir2.1 mRNA的表达水平,发现再灌注后Cx43 mRNA同样发生明显下降,而kir2.1 mRNA则未发生明显变化。这提示再灌注后Cx43的表达除了受到miR-1存在于转录后水平的调节,在转录水平还受到其他因素影响其表达,而缺血/再灌注过程β肾上腺素能受体信号通路的过度激活可能就是影响因素之一[15]。
综上所述,内源性N/OFQ能够上调miR-1,抑制心肌细胞Cx43及kir2.1的表达,进而导致RA。本研究发现了内源性孤啡肽受体信号通路调节心肌损伤过程中的微小RNA机制,能够为其他感觉神经肽以及经典阿片受体系统调控心肌损伤与保护提供新的思路。
(致谢:本实验在山西医科大学第二医院麻醉实验室完成,感谢梁蔚骏老师的支持和帮助,也感谢实验室各位同学的指导与帮助!)
猜你喜欢
中国心脏起搏与心电生理杂志(2023年6期)2024-01-04 12:10:24
人人健康(2022年14期)2022-07-26 02:29:08
世界科学技术-中医药现代化(2022年2期)2022-05-25 13:16:04
世界科学技术-中医药现代化(2021年7期)2021-11-04 08:10:24
实用心电学杂志(2021年5期)2021-10-25 08:34:28
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
实用心电学杂志(2020年6期)2021-01-13 06:31:48
科学(2020年3期)2020-11-26 08:18:34
海南医学(2016年8期)2016-06-08 05:43:00
中国病理生理杂志(2015年8期)2015-12-21 12:38:08
