
脑红蛋白在脊髓损伤后星型胶质细胞内的表达及意义
2019-03-08 00:44明超曾云熊敏
山东医药 2019年4期
明超,曾云,熊敏
(湖北医药学院附属东风总医院,湖北十堰 442000)
脊髓损伤(SCI)分为原发性和继发性,原发性SCI以神经细胞坏死和神经纤维断裂、血-脊髓屏障破坏、血管损伤等表现为主[1];继发性SCI以细胞凋亡和进展性轴突丢失为主,主要包括微循环缺血缺氧、氧化应激损伤、兴奋性氨基酸中毒、炎症反应等[2]。研究表明氧化应激损伤在继发性SCI的发生发展中发挥重要作用[3]。氧自由基(ROS)的堆积会造成脊髓组织氧化应激损伤,主要包括膜脂质过氧化反应、核酸和蛋白质损伤、氧化呼吸链酶系统抑制、神经纤维退变及脱髓鞘等,最终导致脊髓功能紊乱和损伤进一步加重[4,5]。星形胶质细胞(AS)是中枢神经系统(CNS)内分布最广、数量最多的一类细胞。研究表明AS可通过分泌神经营养因子、减轻氧化应激损伤及缺血缺氧预适应等机制保护神经元[6]。SCI后星形胶质瘢痕可减轻神经炎症反应,抑制轴突及神经纤维脱髓鞘而保护神经细胞免受二次损伤[7]。Anderson等[8]提出胶质瘢痕有助于SCI后轴突再生。研究认为,SCI后AS的活化是保护脊髓组织功能及修复的潜在靶点[9]。Ngb是一种定位于细胞质的携氧球蛋白,主要分布于神经系统、视网膜细胞及内分泌腺组织中[10]。研究发现Ngb具有清除氧化应激毒性物质、调控凋亡通路、激活细胞生存信号等功能[11]。在脑损伤、脑疟疾及自身免疫性脑炎中,Ngb除了表达于神经元,亦能表达于损伤区域的反应性AS中;且与星形胶质瘢痕形成有关[12,13]。Ngb在SCI后对神经元的保护作用已有诸多报道,而在AS中的作用却未见确切报道。2017年1月~2018年2月,我们拟从动物模型上探讨SCI后AS中Ngb的表达变化,在细胞模型中研究Ngb在保护AS免受氧化应激损伤中的作用。
1 材料与方法
1.1 材料 雌性SD大鼠30只(3月龄,体质量250~300 g)、SD新生乳鼠10只由湖北医药学院实验动物中心提供。主要试剂:H2O2(美国Sigma公司);兔抗大鼠GFAP及小鼠抗大鼠Ngb抗体(英国Abcam公司);驴抗兔Cy3及驴抗小鼠FITC(武汉Antgene公司);胎牛血清FBS(以色列BI公司);DMEM高糖培养基(美国Thermo公司);TRIzol试剂(美国MRC公司);PCR引物(上海生工生物有限公司设计合成);逆转录试剂盒(美国Fermentas公司)。主要仪器:IX71 荧光显微镜(日本Olympus公司);CO2细胞培养箱(美国Thermo公司);FACS流式细胞仪(美国BD公司);全波段酶标仪(美国Biotek公司);荧光定量PCR仪(澳大利亚RotorGene公司)。
1.2 SCI模型制作 将30只SD大鼠随机分成2组,假手术组(5只)及SCI组(25只)。SCI组采用改良Allen′s 法建立脊髓损伤模型[14],10%水合氯醛(350 mg/kg)麻醉大鼠,俯卧固定脱毛,以浮肋与第13胸椎连接处作为定位标志,后正中线纵行3 cm切口,依次暴露椎旁软组织,咬除棘突、椎板,显露 T10脊髓。脊髓冲击棒(10 g)于25 mm处垂直自由下落撞击T10硬脊膜,持续压迫约10 s,造成中度脊髓挫伤,分层缝合后分笼喂养。SCI成功标准:脊髓组织出血,硬脊膜膨隆呈紫红色,鼠尾痉挛性摆动,双下肢及躯体回缩样扑动,呈迟缓性瘫痪。本实验均获湖北医药学院动物研究伦理委员会批准。
1.3 脊髓组织取材及HE染色 术后第1、3、7、14、28天分别于SCI组随机选取5只大鼠及假手术组选取3只大鼠,麻醉后暴露心脏,经升主动脉插管灌注生理盐水250 mL,待右心耳流出无色清亮液体后继以4% 多聚甲醛溶液灌注,随后打开椎管,分离出损伤处脊髓,4%多聚甲醛固定24 h后,石蜡包埋切片(4 μm)行HE染色。在SCI后1 h取出脊髓组织,可见其连续性中断、表面严重变性坏死,损伤周围亦有不同程度的变性坏死,假手术组大鼠脊髓完整,未见明显出血坏死。
1.4 脊髓星型胶质细胞取材及鉴定 取1~2 d的SD新生乳鼠,经75%乙醇浸泡10 min后处死,超净台中取出脊髓,剥离脊膜及血管,置于冰PBS溶液中剪碎,0.25%胰酶溶液室温消化15 min,12%FBS-DMEM终止消化制成细胞悬液。200目滤网过滤、离心、重悬后接种至T25培养瓶。以差速贴壁法去除成纤维细胞。24 h后换液,以后每隔2~3 d换液1次。取6~8 d培养至90%融合的细胞于37 ℃恒温摇床中160 r/min水平振荡,16~18 h去除少突胶质细胞及小胶质细胞。待细胞传至第三代时(约21 d),取少量细胞以GFAP及DAPI双染细胞免疫荧光法鉴定AS及其纯度。在染色视野范围内,可见GFAP阳性细胞胞体突起之间排列紧密,GFAP和DAPI共表达的细胞占有核细胞总数的90%以上,故而选取第三代AS用于后续实验。
1.5 GFAP及Ngb免疫荧光染色 第三代AS以1×105/mL密度接种于明胶包被的24孔板中,待细胞融合至80%左右时预冷PBS漂洗3次,4%多聚甲醛4 ℃固定30 min,PBS漂洗5 min×3次,后续步骤同脊髓组织免疫荧光染色。免疫荧光染色观察AS内GFAP和Ngb表达。
1.6 AS细胞活力检测 采用MTT法。第三代AS以5×104/mL密度接种于96孔板中,每孔200 μL,置于CO2培养箱中培养72 h。待细胞融合至80%后,分别加入不同浓度(0、50、100、150、200、400 μmol/L)H2O2处理不同时间(6、12 h)用于MTT实验。全波段酶标仪测量波长490 nm时各孔吸光度(A)值。细胞活力=(药物组A值-调零孔A值)/(对照孔A值-调零孔A值)×100%。
1.7 AS内源性ROS生成测定 采用流式细胞术。第三代AS按1.25×105/mL密度接种于6孔板中,每孔2 mL,细胞培养箱中培养72 h后,不同浓度(50、100、200、400 μmol/L)H2O2处理不同时间(6、12 h),胰酶消化、重悬后加入终浓度为10 μmol/L的DCFH-DA探针,37 ℃孵育1 h。无血清培养基洗涤、离心后,2%FBS-DMEM重悬细胞,流式细胞仪检测。
1.8 Ngb、SOD2、CAT mRNA表达检测 采用实时荧光定量PCR法。取不同浓度(50、100、200、400 μmol/L)H2O2处理不同时间(6、12 h)后的AS,TRIzol法抽提总mRNA,应用逆转录试剂盒将1 μg总RNA逆转录成cDNA,稀释至5 ng/μL,随后加入SYBR Green PCR Master Mix及相应引物进行荧光定量PCR反应。引物序列分别为:Ngb正向引物5′-GGCCATCCAAGAGAGGTGAT -3′,Ngb反向引物5′-CCATGCCTCCTCACTACCAA-3′;SOD2正向引物5′- CTGCTGGGGATTGATGTGTG-3′,SOD2反向引物5′-CTACAAAACACCCACCACGGCAT-3′;CAT正向引物5′-GCGGATTCCTGAGAGAGTGG-3′,CAT反向引物5′- GAATCGGACGGCAATAGGAG-3′;反应条件:95 ℃ 2 min,95 ℃ 5 s,60 ℃ 30 s,72 ℃ 30 s,72 ℃ 5 min共35个循环。以2ΔΔCt法计算目的基因相对表达量。
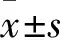
2 结果
2.1 SCI后AS内GFAP及Ngb表达 假手术组脊髓组织内少量Ngb表达,而GFAP阳性细胞呈散在分布,白质内AS胞体小,胞突细长且直,分支较少。在脊髓边缘的白质内可见极少量GFAP和Ngb共表达于细胞质中,且显色很弱。SCI组术后1 d,可见脊髓组织结构破坏严重,损伤区域周围Ngb阳性细胞数目开始增加,与假手术组相比,GFAP和Ngb共表达稍有增加。术后3 d,Ngb表达继续增加,损伤区域周围GFAP和Ngb共表达明显增加;术后1周,GFAP表达开始显著增加,并聚集于损伤区域边缘。与之相似,GFAP和Ngb共表达亦显著增加,明显高于临近未损伤区域;术后14 d及28 d内,GFAP的增长速度最快,28 d时达到峰值,此时AS胞体突起增多、延长,突起之间形成致密网络,形成囊腔与空洞的边界,而周围区域GFAP和Ngb共表达却在14 d时达到峰值,28 d时共表达减弱。
2.2 H2O2不同浓度及时间处理AS细胞活力变化 见表1。H2O2处理6 h或12 h后,随着H2O2浓度的增加,AS活力逐渐降低(F6 h=137.36,P<0.01;F12 h=143.61,P<0.001)。H2O2处理6 h后,100与150 μmol/L间AS活力无统计学差异(P=0.158);与之相似,12 h后150与200 μmol/L间亦无统计学差异(P=0.168)。因此确立H2O2浓度为50、100、200、400 μmol/L用于后续实验。随暴露时间延长,同一浓度下AS活力持续降低,而50 μmol/L H2O2时却无统计学差异。

表1 H2O2不同浓度及时间处理AS活力变化
注:6 h时与H2O2未处理细胞相比,*P<0.01;12 h时与H2O2未处理细胞相比,#P<0.001。
2.3 H2O2处理6、12 h诱导ROS生成情况 见图1及图2。H2O2可诱导AS产生ROS,且在6 h内,随着H2O2浓度的增加,ROS生成逐渐增加(峰值右偏),差异有统计学意义(F=262.33,P<0.01)。暴露于H2O212 h的AS,ROS生成先升后降(F=297.30,P<0.01),并在100 μmol/L时达到峰值。

图1 H2O2处理6 h诱导ROS生成情况

图2 H2O2处理12 h诱导ROS生成情况
2.4 不同时点H2O2诱导Ngb、SOD2、CAT mRNA表达变化 见图3A、B、C。暴露于50~400 μmol/L H2O26 h或12 h后,Ngb mRNA表达呈先升后降趋势(F6 h=67.48,P<0.01;F12 h=51.47,P<0.01)。
且在100 μmol/L 6 h或12 h时均达到峰值(t=4.72,P=0.042)。与之相似,CAT、SOD2mRNA表达亦呈先升后降趋势(CAT:F6 h=95.31,P<0.001;F12 h=101.20,P<0.001,SOD2:F6 h=94.73,P<0.001;F12 h=41.54,P<0.01),而暴露于100 μmol/L H2O212 h时,SOD2表达量高于6 h(t=6.78,P=0.032)。Ngb、CAT、SOD2mRNA表达在50 μmol/L或100 μmol/L时达到峰值,随后持续降低,400 μmol/L时达到最低值,呈现剂量-时间依赖效应(P均<0.05)。

图3 H2O2处理AS抗氧化应激基因表达
3 讨论
氧化应激损伤是促氧化剂和抗氧化剂平衡失调、自由基产生过多、机体或组织倾向于氧化,导致中性粒细胞浸润、蛋白酶分泌增加,产生大量氧化中间产物的一种状态。Bermudez等[15]研究发现,氧化应激在SCI的急、慢性期均持续存在。Amri等[19]研究发现,100 μmol/L的H2O2可使大脑皮层AS发生皱缩、胶质网络发生塌陷、细胞密度减少;然而加入Ngb后可阻止这些改变的发生,同时Ngb还可通过减少ROS形成及促炎因子的表达而发挥抗凋亡作用[16]。
哺乳动物CNS的再生能力贫乏,啮齿类动物(完全横断性脊髓损伤除外)在SCI急性期过后脊髓功能却发生一定程度的自然恢复。就CNS损伤后的细胞反应而言,人类进化选择似乎更倾向将小伤害隔离同时预防损伤区域感染,而不是去修复大且无法复原的损伤,AS的行为正好彰显了这一特点。SCI后相比于AS而言,神经元细胞更易受到ROS攻击,AS具有保护神经元免受氧化应激损伤的作用。一方面与AS含有更多的谷胱甘肽及维生素E等抗氧化物质有关,另一方面亦与细胞进化的自然选择有关。
研究表明SCI能触发AS增殖与活化,而活化的AS可抑制SCI区域的扩大、清除细胞碎片、重塑损伤区域[17]。Nathan等[18]研究结果显示,脊髓环境可改变AS的命运。Okada等[7]研究证实在SCI恢复过程中,敲除AS内Socs3活化抑制基因后,AS迁移和隔离损伤区域及限制炎症播散的能力均增强。本研究证实,在SCI的自然恢复过程中,SCI可促进AS中Ngb表达。SCI早期Ngb表达增加可能与其增加损伤区域的氧供有关。在SCI后7 d,损伤区域周围GFAP和Ngb共表达开始明显增加,14 d达到峰值,此时胶质瘢痕形成的速度最快,Ngb的这种变化趋势与AS功能相匹配,由此推断Ngb可能促进了AS的活化,从而促进SCI的自然恢复。而SCI后1~28 d Ngb呈现先升后降的表达趋势,说明除了AS的活化,Ngb还可能扮演抗氧化应激损伤的作用,此与Amri等[16]及Fabrizius等[19]研究结果一致。
ROS是反映氧化应激的指标。本实验结果显示,不同浓度H2O2处理脊髓AS 6 或12 h后,细胞出现不同程度的氧化应激损伤,AS活力逐渐降低,呈现剂量-时间依赖效应。实时荧光定量PCR结果显示,H2O2诱导AS氧化应激损伤后可促进SOD2、CAT、Ngb基因表达;且随着H2O2浓度增加及暴露时间延长,内源性抗氧化应激酶SOD2及CAT mRNA水平呈现先升后降趋势。与此同时,Ngb mRNA亦出现类似于SOD2、CAT基因的表达趋势,且在100 μmol/L时达到峰值。说明Ngb可能发挥类似作用,与SOD2、CAT共同参与AS的抗氧化应激损伤。Douiri等[20]将外源性Ngb加入H2O2诱导损伤后的AS中发现,Ngb可明显增加SOD2及CAT mRNA表达,从而发挥抗氧化应激损伤作用;而抑制AS内源性抗氧化酶系统的活性则可明显削弱内源性保护分子促进细胞存活的能力。
本研究结果证实SCI可促进AS表达Ngb,并且Ngb可能参与了SCI后AS的抗氧化应激损伤作用,从而促进SCI的自然恢复过程。此为星型胶质细胞与脑红蛋白抗氧化应激损伤治疗脊髓损伤奠定了理论基础。
猜你喜欢
军事文摘(2022年8期)2022-11-03
安徽农业科学(2022年1期)2022-02-14
江苏农业学报(2021年2期)2021-06-30
昆明医科大学学报(2020年12期)2021-01-26
神经损伤与功能重建(2020年10期)2020-12-23
神经损伤与功能重建(2020年11期)2020-12-01
神经损伤与功能重建(2018年2期)2018-02-01
湖北农业科学(2017年10期)2017-06-22
中国运动医学杂志(2016年3期)2016-07-10
中国生化药物杂志(2015年4期)2015-07-07
